
А. С. Раутиан палеонтология как источник сведений о закон
| Вид материала | Закон |
- Культурное разнообразие в обществе, институты и экономическое развитие: вопросы теории, 129.28kb.
- Конституция Российской Федерации: проблемы реализации. Толкование Конституции России., 21.07kb.
- Программа курса лекций, 64.32kb.
- Современный урок истории немыслим без документов, исторических первоисточников, произведений, 71.78kb.
- Современный урок истории немыслим без документов, исторических первоисточников, произведений, 70.47kb.
- А. С. Раутиан Самое замечательное, что может быть в человеческой жизни, 792.34kb.
- Федеральный закон, 375.18kb.
- "Простите вы нас, родители. Может, мы вас обидели ". Источник: Региональный еженедельник, 176.14kb.
- Конституция Российской Федерации как источник правового обеспечения управленческой, 41.89kb.
- Конституция Российской Федерации как источник уголовно-процессуального права. 13., 120.76kb.
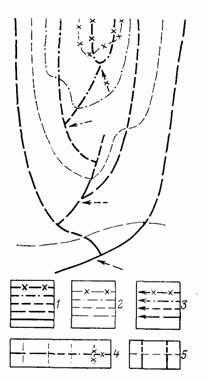
Рис. 15. Схема филогенеза в соответствии с концепцией аристогенеза. Составил А. С. Раутиан. 1 — филетические линии; 2 — границы уровней организации; 3 — моменты приобретения ключевых ароморфозов вышестоящих уровней организации; ряды Э. Д. Копа: 4 — гомологические, 5 — гетерологические
Можно указать направления специализации, характерные для всех стволов животного царства, удовлетворяющих правилу прогрессивного совершенствования [1130, 1382]. Эти направления отражены в следующих правилах: прогрессивной дифференциации, интеграции и разделения функций [1192, 1196, 1382]; автономиза-ции и рационализации онтогенеза [1192, 1196, 1197]; цефализации — увеличения относительных размеров и концентрации центральной нервной системы [1919]; возрастания активности и чувствительности [250].
Сравним особенности прогрессивной специализации и дестабилизации (эманации) как основных модусов эволюции:
Прогрессивная специализация Дестабилизация (эманация)
Соотношение запоминания и забвения Запоминание превосходит забвение Забвение превосходит запоминание
Надежность запоминания Возрастет Понижается
Объем памяти Возрастает Понижается
Преемственность (надежность наследования) Возрастает Понижается
Устойчивость Возрастает Понижается
Свобода выбора (изменчивости) Понижается Возрастает
Типичная длительность Продолжительна Очень коротка
Корреляции в онтогенезе Возникают Разрушаются
Таксономический уровень изменчивости признаков Понижается Возрастает
Степень приспособленности (адаптированность) Возрастает Понижается
Направленность эволюции Возрастает Понижается
Предел развития Персистирование, или переход к дестаби- Разрушение структуры (вымирание), или
лизации (эманации) переход к специализации
Третий из качественных режимов (модусов) развития (см. рис. 9), как отмечалось, является переходным и наблюдается в момент смены одного из двух основных модусов эволюции другим.
Онтогенез как прогрессивная специализация. Ярчайшим проявлением параллельной и итеративной специализации является повторяемость онтогенетических процессов — индивидуальной специализации организмов каждого вида. В высшем проявлении онтогенез многоклеточных абсолютно необратим и сопровождается программируемой смертью. У простейших онтогенез обратим, а особь потенциально бессмертна. Строго не ограниченные определенной стадией жизненного цикла процессы генетического обмена у прокариот принципиально допускают существование прокариотического бионта, развивающегося в разные моменты жизни на базе разных геномов. По этому важнейшему признаку у них оказывается неопределенной грань между индивидуальным и историческим развитием. Перечисленные типы организации (прокариоты — простейшие — многоклеточные) могут рассматриваться как узловые стадии разделения процессов онто- и филогенеза (рис. 16). Биологический смысл их разделения заключается в снятии противоречия между высокой организацией, требующей для своего осуществления высокого уровня устойчивости и автономности [1192, 1196], и требованием высокой лабильности, свободы творчества для продолжения развития. Снятие это осуществлено путем разделения проявлений жизни на популяционный и организменный уровни организации.
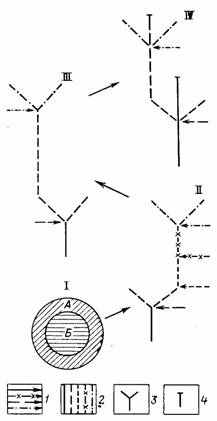
Рис. 16. Узловые стадии становления типичного онтогенеза метазоа и метафита как процесса тотальной специализации. Составил А. С. Раутиан. I — гипотетическая стадия разделения сомы (А) и гермы (Б); схемы изображения онтогенезов: II — прокариотического организма; III—эукариотического простейшего; IV — метазоа и метафита; 1 — вставки чуждой гермы в процессы онтогенеза; 2 — фазы онтогенеза, осуществляющиеся без вмешательства новой гермы; 3 — размножение; 4 — смерть
Популяция — низко организованная система с относительно слабыми связями между гомономно построенными, автономными и взаимозаменимыми элементами (особями), способная существовать продолжительное время, но медленно и обратимо специализирующаяся (обучающаяся), оказывается способной к эволюции. Организм—высокоорганизованная система с весьма сильными связями между гетерономно построенными, уникальными (незаменимыми), но подчиненными целому элементами (органеллами, органами, тканями), имеющая ограниченный срок существования вследствие совершенства памяти и необратимости процессов быстрого обучения (специализации). Сопряжение между обоими уровнями осуществляется благодаря процессу размножения, а обратная связь—благодаря процессам отбора (выбора) [1192, 1193, 1196]. Цена снятия отмеченного противоречия—регулярная эманация индивидуальной организации и опыта в процессах размножения, а в своих высших формах—смерть.
Правило специализации экто- и эндосоматических подсистем организма в филогенезе. Адаптация как выражение прогрессивной специализации носит характер прогрессивной оптимизации целого. Пути этой оптимизации сталкиваются с ограничением: оптимизация целостной системы по каждому параметру осуществляется в ущерб оптимизации по любому другому параметру в меру их взаимной сопряженности [754, 859, 1223]. Поэтому процесс специализации осуществляется тем легче и быстрее, чем меньшее число противоречивых структурно-функциональных подсистем в него вовлечено. Действительно, обычно процесс специализации удается особенно тщательно проследить на примере отдельных функциональных подсистем [19—21], например опорно-двигательного аппарата [215, 495, 1023].
Число главных функций, на которые делается адаптивная ставка, всегда невелико. Все прочие эволюируют в координационном порядке, обслуживая в конечном счете оптимизацию главных. Следуя духу, но не букве определений А. Н. Северцова [926], главные структурно-функциональные подсистемы целого можно назвать эктосоматическими, а подчиненные—эндосоматическими. Соответственно семофилезы (филогенетические изменения части или аспекта целого) первых можно назвать проталлаксами, а вторых — дейталлаксами.
Сравним особенности двух основных классов функциональных систем организма, отличающихся по характеру семофилеза в процессе прогрессивной специализации и роли в обеспечении адаптации целого организма:
Эктосоматические системы Эндосоматические системы
Типы семофилезов, осуществляющихся в процессе специализации
Проталлаксы Дейталлаксы
Количество в организме
Малое Очень большое
Роль в процессе адаптации к внешней среде
Возрастающая Понижающаяся
Роль в процессе коадаптации частей целого
Понижающаяся Возрастающая
Вклад в достигнутую приспособленность организма
Преимущественно — устойчивость к внешней Преимущественно — координация
среде частей и поддержание целостности
Внутренняя дифференцировка функциональной системы
Прогрессивная Ограниченная, распространены редукции
Семофилез отношений к организму, как части к целому
Прогрессивная эмансипация в рамках целого Прогрессивное подчинение целому
организма организму
Роль в стратегии эволюции организма
Определяющая Компенсаторная
Роль в поддержании устойчивости
Поддержание направления специализации — Поддержание каждого состояния
гомеорезис филогенеза в их последовательном ряду —
гомеостазис таксона
Наиболее характерные формы специализация принимает, когда значительный и долгосрочный прогресс устойчивости (адаптации) достигается при участии минимума эктосоматических подсистем. При этом прогресс будет достигаться при минимуме новизны и опасности вымирания, а направление специализации будет в максимальной степени сохраняться.
Эктосоматические подсистемы по мере их прогрессивной специализации все в возрастающей степени берут на себя функции поддержания устойчивости целого во внешней среде. Это сопровождается дифференцировкой, ростом индивидуальности и эмансипированности в рамках целого. Специализация эктосоматических подсистем достигается ценой прогрессивного подчинения им специализации эндосоматических подсистем и ограничения возможностей независимой от них специализации последних. Такой путь ведет к нарушению целостности (понижению адекватности внутреннего умвельта), причем, как ни странно, в результате развития, направленного на достижение все большей устойчивости целого. Относительно бесконфликтный, направленный и безопасный путь специализации ведет к накоплению внутренних противоречий устойчивости (достигнутого уровня адаптации) и лабильности (свободы выбора и перспективы адаптиогенеза). Эти противоречия сначала останавливают, а затем и разрушают данное направление специализации. Основные фазы процесса прогрессивной специализации приведены на рис. 10. Пример такого процесса, исследованного Т. Уэстоллом [2482] для двоякодышащих рыб, показан на рис. 17.
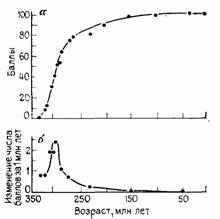
Рис. 17. Специализация и темп аристогенеза двоякодышащих рыб от силура (по реконструкции) до наших дней. По Т. С. Уэстоллу. а — уровень специализации — степень обновленности признаков (преобразования в направлении их современных модальностей), выраженная в условных средних баллах; б—темп аристогенеза—скорость обновления признаков, осуществлявшегося в нескольких параллельно развивавшихся филетических линиях
Правило адаптивной редукции. Оптимизация (адаптация) целого возможна не только за счет дифференцировки и усложнения структуры, при структурном превосходстве системы над средой, но и за счет упрощения структуры—сокращения затрат на поддержание частей (аспектов), функциональное значение которых в поддержании устойчивости не оправдывает себя. Возрастающая роль эктосоматических подсистем по мере специализации понижает относительную адаптивную ценность многих эндосоматических подсистем, для которых и характерны явления адаптивной редукции. В условиях специализации морфофизиоло-гический прогресс протекает параллельно с редукцией; эти процессы как минимум компенсируют друг друга (телогенез) или даже редукция опережает морфофизиологический процесс (катагенез, гипогенез, рационализация онтогенеза [1197]).
Разнообразные формы вторичного упрощения и односторонний морфофизиологический прогресс немногих эктосоматических подсистем (наравне с эволюционными тупиками и вымиранием)—весьма характерные спутники специализации [805, 935, 1197]. Поэтому указания на роль специализации в процессах ароморфной эволюции появились лишь в самое последнее время [933—936], хотя, как мы уже знаем, общий рост уровня организации в конечном счете должен достигаться именно вследствие процессов специализации.
Правило мозаичной эволюции. Данное правило свидетельствует о широком распространении в филогенезе неравномерностей темпов специализации разных структурно-функциональных подсистем целого [594, 935, 1033, 1040]. Статическим его аналогом является правило гетеробатмии — разной эволюционной продвинутости разных частей целого [935, 936, 1040], а онтогенетическим — правило исторического гетерогенезиса стадий индивидуального развития [540, 887].
Эти правила обычно интерпретируются как свидетельство значительной свободы и независимости поведения отдельных признаков в филогенезе [594,596], принадлежащих разным координационным цепям [935, 936, 1040], низкой целостности филогенетических систем [2046, 2048, 2053], а следовательно, низкой преемственности их состояний как целого. В пользу такой позиции говорит лишь широкое распространение и наглядность явлений мозаичности при становлении ароморфных групп высокого ранга [1033], которому должна предшествовать глубокая эманация, а значит, и понижение целостности. Однако само становление есть выражение специализации — повышения устойчивости целого и взаимозависимости его частей. Эволюционная разноуровневость подсистем целого при гетеробатмии [935, 936, 1040] является следствием несинхронного появления признаков нарождающегося таксона. При этом организация переходных групп обычно совмещает признаки предкового и дочернего таксонов с чертами собственной специализации. Реальные предковые формы лишь в редких случаях бывают промежуточными по организации, как это обычно полагалось до начала палеонтологической эры в филогенетике [1033, 1034]. Сама возможность поляризации признаков по уровням говорит о направленности и преемственности несинхронных, но взаимообусловленных семофилезов целого.
Особенно ярко противоречие мозаичности и ортогенетической направленности эволюции выразилось в концепции аристогенеза Г. Осборна [291, 2046, 2048, 2053]. Однако все эти факты приходят в согласие, если предположить, что главной причиной мозаичности является запрет на синхронную оптимизацию многих разноплановых эктосоматических подсистем целого и большие порции кратковременно получаемой новизны, которых требует такая синхронная оптимизация. Это хорошо согласуется с мнением А. Л. Тахтаджяна, что при освоении новой адаптивной зоны меняются в первую очередь те подсистемы, которые определяют адаптацию к новым условиям; остальные подсистемы отстают в своем филогенетическом развитии [935, 936, 1040]. При этом эффект мозаичной эволюции представляет собой следствие не мозаичной организации, а, напротив, целостности филогенетической системы и согласованности составляющих его семофилезов. Таким образом, мозаичность эволюции — следствие комплексной оптимизации целого, которая возможна только как процесс последовательной (несинхронной) оптимизации отдельных его блоков, смены ролей экто- и эндосоматических подсистем, а значит, и смены направлений специализации.
Правило инадаптивной специализации. Правило отражает необходимую односторонность и противоречивость специализации целого. Накапливающиеся в этом процессе противоречия целого и частей (нарушение адекватности внутреннего умвельта), устойчивости и способности к изменению (ограничения свободы творчества) и т. п. в конечном счете начинают тормозить дальнейшую прогрессивную специализацию в данном направлении, затрудняя смену ее направления, ставя развивающуюся систему под угрозу гибели. Все эти оборотные стороны специализации могут рассматриваться как инадаптивные ее последствия, «плата» за приобретенную устойчивость — адаптацию [495].
Инадаптивная специализация, таким образом, не особая форма специализации, а термин для обозначения одного из имманентно присущих ей свойств. Интерпретация особенностей организации одного таксона как инадаптивных по сравнению с другим позволяет объяснить причину явления конкурентного вытеснения, биологического регресса инадаптивного таксона на фоне процветания экологически близкого, но эвадаптивного, по терминологии А. П. Расницына, таксона [859]. Экологически близкие таксоны. достигшие сравнимого уровня специализации, могут отличаться по инадаптивным последствиям и, следовательно, перспективам будущей специализации.
Д. Роза обратил внимание на относительную «скороспелость» инадаптивных и «запоздалость» эвадаптивных таксонов [291]. Действительно, тем легче и быстрее протекает прогрессивная специализация, а инадаптивность приобретает критические размеры, чем более односторонен этот процесс, чем меньшее число эктосоматических подсистем его определяет, чем более они индивидуальны и чем в большей степени определяют судьбу эндосоматических подсистем целого. «Запоздалые» таксоны, напротив, благодаря низкому темпу специализации могут обеспечить более гармоническое развитие целого с участием большего числа экто-соматических подсистем и более полной компенсацией их одностороннего развития эндосоматическими подсистемами, т. е. позволить менее противоречивое и более компромиссное развитие функциональных систем целого [859]. В процессе филогенеза крупного таксона первым биологического прогресса достигает обычно наиболее инадаптивный подтаксон, который вскоре последовательно сменяют все более эвадаптивные подтаксоны. Наличие двух таких смен Д. Роза считал правилом.
Правило итеративной эволюции. Если некоторое направление прогрессивной специализации исчерпало себя вследствие накопления груза инадаптивности (что завершается той или иной формой биологического регресса), то в сходном направлении может возникнуть новый цикл прогрессивной специализации, начальные этапы которого, естественно, будут уступать по уровню специализации конечным звеньям предыдущего цикла. Такой повторяющийся во времени процесс прогрессивной специализации в очень близком направлении, начинающийся каждый раз от родственных, часто персистирующих, но слабоспециализированных в данном направлении форм, составляет содержание итеративной эволюции [864]. Иногда итерация (возврат) осуществляется путем вторичной деспециализации (например, путем феталнзации) ина-даптивного филума [248]. Итеративная эволюция — особый случай широко понимаемого явления параллелизма. Повторение очень близких направлений специализации свидетельствует о большой специализации предка итеративных филогенезов, что хорошо согласуется с его персистированием. Сами итеративные филогенезы чаще всего являются гиперморфными линиями, завершающимися вымиранием или реже фетализацией, но не персистированием. Итеративная эволюция показывает, что порой проще создать новое, сходное с прежним, направление специализации, чем избавиться от накопленного груза инадаптивности.
Примеры итеративной эволюции известны в филогенезе двустворчатых моллюсков [24], аммонитов [887, 864], акул [248] и других групп.
Правило адаптивной радиации. Стабилизация новообразования на каждом уровне рассмотрения по отношению к итогу прежней стабилизации представляет собой в большей или меньшей степени смену направления специализации. Это хорошо согласуется с представлением И. И. Шмальгаузена об аллогенезе [805, 1197], при котором освоение таксоном новой адаптивной зоны начинается с одной или немногих субзон [952]. Напротив, при разделении первоначально широкой адаптивной зоны предка на субзоны [952] причиной адаптивной радиации является специализация каждого из подтаксонов, достигающаяся путем интенсивной эксплуатации части ресурсов адаптивной зоны предка. Эти два способа адаптивной радиации нельзя считать альтернативными: любая специализация до известной степени изменяет адаптивную зону, а любая смена адаптивных зон немыслима вне специализации.
Однако в пределах заданного направления прогрессивной специализации таксономический уровень, на котором осуществляется адаптивная радиация, будет убывать пропорционально достигнутому уровню специализации (рис. 18).
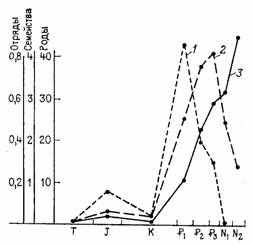
Рис. 18. Падение таксономического уровня, на котором протекала адаптивная радиация млекопитающих, по мере их прогрессивной специализации. По Дж. Г. Симпсону. Скорость появления (число соответствующих таксонов за 1 млн. лет—отложено по ординате); 1 — отрядов, 2 — семейств, 3 — родов
После эманации, если не наступает вымирание, прежде всего происходит стабилизация структурного типа нового таксона, останавливающая дальнейшую эманацию. На базе еще достаточно широкого спектра изменчивости, но уже более совершенного механизма запоминания новообразований активизируется адаптивная радиация. Поскольку устойчивость нового таксона еще невелика, большинство ради-ирующих ветвей окажутся недолговечными. Более продолжительную историю будут иметь ветви, продолжающие процесс специализации в избранном направлении, а значит, приобретающие все большую ограниченность и устойчивость (адаптацию). Именно этот процесс, первоначально расчленяющий единую адаптивную зону на субзоны, со временем начинает постепенно замедлять дальнейшую адаптивную радиацию на данном таксономическом уровне и все больше стимулировать ее на следующем, более низком (см. рис. 9). Возрастающая устойчивость предыдущего иерархического уровня будет оказывать все более канализирующее воздействие на эволюцию нижележащего, вводя его в русло прогрессивной специализации.