А. С. Раутиан палеонтология как источник сведений о закон
| Вид материала | Закон |
| Эмпирические закономерности (правила) эволюции Рис. 13. Аристогенез в эволюции териодонтов. По |
- Культурное разнообразие в обществе, институты и экономическое развитие: вопросы теории, 129.28kb.
- Конституция Российской Федерации: проблемы реализации. Толкование Конституции России., 21.07kb.
- Программа курса лекций, 64.32kb.
- Современный урок истории немыслим без документов, исторических первоисточников, произведений, 71.78kb.
- Современный урок истории немыслим без документов, исторических первоисточников, произведений, 70.47kb.
- А. С. Раутиан Самое замечательное, что может быть в человеческой жизни, 792.34kb.
- Федеральный закон, 375.18kb.
- "Простите вы нас, родители. Может, мы вас обидели ". Источник: Региональный еженедельник, 176.14kb.
- Конституция Российской Федерации как источник правового обеспечения управленческой, 41.89kb.
- Конституция Российской Федерации как источник уголовно-процессуального права. 13., 120.76kb.
ЭМПИРИЧЕСКИЕ ЗАКОНОМЕРНОСТИ (ПРАВИЛА) ЭВОЛЮЦИИ
Правило направленности эволюции (ортогенеза). Пока действует заключенная в структуре система запретов, вся динамика развивающейся системы будет обладать свойством гомеорезиса — направленного канализованного развития. Развитие не может быть абсолютно ненаправленным.
Между двумя состояниями системы, предыдущим и последующим, можно указать вектор изменения. Такая направленность касается только данного акта выбора и в общем случае не требует памяти. Указание направления каждого акта выбора не означает направленного развития. Последнее возникает там, где неслучайные процессы охватывают не менее трех преемственно связанных состояний (предшествующее операнду состояние—операнд— выбор), обозначенных нами как элементарное звено развития. В этом случае состояние, предшествующее операнду, может влиять на выбор только благодаря памяти.
Оператор непосредственно может влиять только на исход данной операции выбора, причем единственным образом: ограничивая пространство выбора, заданное структурой операнда. Следовательно, направление развития в рамках данной операции выбора определяется исключительно внутренними свойствами развивающейся системы—операнда. Внешняя среда—оператор— может влиять не только на направление данного, но и последующих выборов, направлять развитие только в меру его отображения в структуре системы—запоминания. Таким образом, роль среды в определении направления развития, как отмечал Ч. Дарвин, исключительно косвенна. Характер и адекватность отображаемого аспекта среды определяется свойствами развивающейся системы. Опосредование оператора операндом в процессе его отображения всегда глубже, чем опосредованно операнда оператором. В противном случае среда реорганизует систему по своему образу и подобию, что равносильно потере индивидуальности и гибели системы. Все рассуждения о прямом влиянии среды на направление биологического развития, а тем более определяющей ее роли, основаны на отсутствии полного разграничения направленности отдельного выбора и направленности развития.
Правило адаптивной направленности развития (адаптиогенеза). Приспособление в биологии часто определяется как ансамбль свойств организации, обеспечивающих устойчивое поддержание в заданных пределах ее количественных и качественных параметров. И это явление можно описать при помощи понятия «устойчивость». Адаптация — понятие, призванное оттенить еще один аспект структуры — конкретные способы поддержания ее устойчивости. Поскольку для структуры альтернатива устойчивости — небытие, всякое длительное развитие должно протекать с явным преобладанием специализации над эманацией, запоминания над забвением, преемственности над новизной и адаптации над процессами ее разрушения. Адаптация — необходимый аспект любой специализации, а ее разрушение — необходимое следствие любой эманации. Это правило накладывает дополнительное граничное условие на предыдущее.
Жизнь, как всякая неравновесная диссипатпвпая структура, поддерживающая свою устойчивость благодаря потреблению внешней энергии и (или) вещества, немыслима без постоянного обмена со средой. Отсюда определение адаптации как соответствия — адекватности структуры системы и среды. Эта адекватность означает наличие структуры отношений системы к среде — умвельта, обеспечивающего поддержание устойчивости системы в среде. Адекватность умвельта важным для жизнедеятельности системы факторам среды и есть адаптация. Можно говорить не только о внешнем, но и внутреннем умвельте—структуре отношений целого к своим элементам. Потеря адекватности внутреннего умвельта фактической структуре элементов равносильна потере целостности.
Возможность активной адаптации для диссипативных, в том числе живых структур, т. е. отыскание ими устойчивых стационарных состояний (приобретение необходимой адекватности ум-вельтов) вдали от состояния термодинамического равновесия, непосредственно вытекает из достижений неравновесной термодинамики и синэргетики.
Адаптация — целостное свойство, аспект структуры. Поэтому из адаптивной стратегии развития целого не следует, что каждая изолированная его часть или аспект должны нести вполне определенную собственную адаптивную функцию. Никакое адаптивное толкование конечного аспекта структуры (указание его роли в поддержании устойчивости целого) не будет полным в меру сопряженности разных аспектов целого и их функций.
Правило постепенности эволюции (градуализма). Постепенное преобразование органических форм в палеонтологических рядах, наличие переходных форм и постепенная трансформация видов из одного в другой в последовательных биозонах были известны еще додарвиновской палеонтологии. Эти факты находились в согласии с представлением об эволюции путем накопления мелких наследственных различий, поэтому Ч. Дарвин рассматривал их как косвенные доводы в пользу своей теории.
Были указаны три граничных условия, при соблюдении которых правило должно выполняться: 1) последовательные биозоны не разделены перерывами в осадконакоплении; 2) в верхней, более молодой, биозоне не происходит замещения близким видом, мигрировавшим из другой географической области; 3) остатки организмов присутствуют в достаточном количестве на протяжении всей осадочной толщи. Иными словами, правило выполняется при условии полноты стратиграфической и адекватности палеонтологической летописи, что почти никогда невозможно утверждать. Эмпирическое обоснование правила на массовом материале летописи практически невозможно. Однако немногие специально-подобранные случаи сравнительно полных стратиграфических и палеонтологических последовательностей с возможно более узкой пространственной локализацией филогенетических процессов позволяют обнаружить тенденцию ископаемых организмов к рядоположенности и гомотаксису с их стратиграфическим положением. Эти «кинематографические» тенденции иногда рассматривают как прямое указание палеонтологии на факт существования эволюции, хотя речь идет о весьма правдоподобной гипотезе.
Постепенность эволюции хорошо согласуется с представлением о разрушительном действии на структуру больших порций кратковременно получаемой новизны. Любые формы макрогенеза сопряжены с большим риском вымирания. Это подтверждает и правило обратной зависимости ранга таксона и частоты возникновения таких таксонов в филогенезе. Постепенное и преемственное развитие, сохраняющее архетип таксона и направление его' специализации, в соответствии с правилом «перемежающегося равновесия» составляет основной фон филогенетических событий. Градуализм нашел отражение в правиле градации или постепенного прогрессивного усовершенствования, правиле постепенного количественного нарастания разнообразия форм жизни от более древних отложений к более молодым. Правило возникновения филогенетических новшеств преимущественно на поздних стадиях онтогенеза — основа «биогенетического закона» Э. Геккеля и закона К. Э. Бэра. Терминальные изменения касаются более мозаичной стадии онтогенеза, в течение которой нарушение одной части меньше вредит развитию других частей целого. Напротив, даже небольшое, но не регулируемое к норме уклонение ранних стадий развития влечет вследствие активного взаимодействия частей на стадиях детерминации цепную реакцию нарушения последующего хода индивидуального развития.
Таким образом, правило постепенности и непрерывности эволюции — просто внешнее выражение преемственности, необходимое условие развития, и оно должно выполнять эвристическую функцию: чем больше кратковременно получаемой новизны требует филогенетическая гипотеза, чем большего размера скачка требуют ее условия, тем менее при прочих равных условиях эта гипотеза правдоподобна.
Правило редкости переходных форм в палеонтологической летописи. Согласно Ч. Дарвину, это правило призвано объяснить несогласие массовых фактов палеонтологической летописи с правилом постепенности эволюции, которое в палеонтологии базируется на выборочных примерах. Чрезмерное увлечение значительными успехами полевой палеонтологии привело некоторых исследователей к выводу, что летопись стала в основном адекватной филогенезу, а наличие разрывов в палеонтологических рядах свидетельствует в пользу макрогенеза. От столь оптимистических выводов удерживают факты отсутствия или крайней редкости ряда таксонов в ископаемом состоянии на протяжении всего или части их филогенеза, необходимость прибегать к понятию «практически вымершего» таксона [367, 368], а также требование преемственности, известной постепенности, филогенеза.
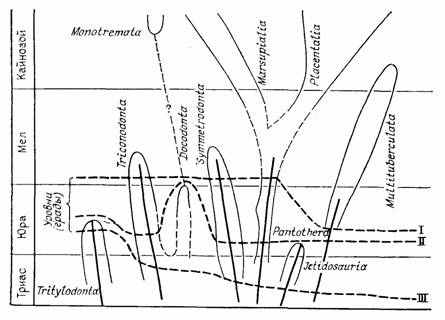
Факт избирательного выпадения именно переходных форм потому и питает идеи макрогенеза, что не может быть объяснен стратиграфической и тафономической неполнотой летописи как таковой. Если существует проблема переходных форм, то, значит, должны быть и продолжительные периоды относительного стасиса таксонов, когда архетип их сохраняется. Для такой фазы филогенеза неоднократно предлагались названия (рис. 10). О. Шиндевольф [2223, 2224] назвал ее типостазом. Чередование длительных периодов стасиса таксонов с относительно краткими периодами дестабилизации [314] является ядром правила «перемежающегося равновесия» [1034, 1550, 1666, 2318]. Вероятность обнаружения группы в палеонтологической летописи, при прочих равных условиях, прямо пропорциональна всем основным критериям биологического прогресса А. Н. Северцова [928]: фактическому таксономическому разнообразию и численности группы, широте ее географического и стратиграфического распространения [457]. Неполнота летописи будет объяснять избирательное выпадение переходных форм, если они приходятся на фазы относительного биологического регресса и сравнительно скоротечны. Об этом говорит даже название «типолиз», данное этой фазе О. Шиндевольфом [2223, 2224]. На эти плохо представленные в летописи моменты филогенеза попадают начала важнейших морфологических перестроек, определяющие становление новых таксонов и смены направлений специализации (типогенез по О. Шиндевольфу). Позиции градуалистов и сторонников прерывистой эволюции отличаются лишь интерпретацией факта редкости переходных форм и, как можно видеть, в чем-то права каждая из сторон.
Правило «перемежающегося равновесия» (пунктуализма). Сформулированное С. Гоулдом, Н. Элдреджем и С. Стенли, это правило гласит: в эволюции видов, как правило, чередуются длительные периоды стасиса, когда основные структурные черты вида сохраняются неизменными, и короткие периоды трансформации одной видовой формы в другую. В основу правила положен не столько массовый материал летописи, хотя он использован тоже, сколько ограниченное число интенсивно исследованных случаев видообразования в стратиграфически и палеонтологически непрерывных или почти непрерывных разрезах. Временные масштабы разрывов оценивались абсолютными методами датирования. С. Гоулд полагает, что этой закономерности подчиняется 95 % случаев видообразования, а С. Стенли оценивает различия в длительности периодов стасиса и трансформации в несколько порядков, измеряя ее предполагаемым числом прошедших поколений [1034]. Авторы связывают свои воззрения с идеей Э. Майра [594, 596] о «бутылочном горлышке» низкой численности, которое проходит популяция в процессе видообразования.
Пунктуализм хорошо согласуется с теорией. Пока система существует, обладает индивидуальностью и выделенностью в среде, ее структура сохраняет большую и меньшую устойчивость (гомеостаз) по отношению к возмущениям. Следовательно, даже непрерывный градиент внешнего воздействия или внутреннего возмущения, по отношению к которому система организована (обладает умвельтом), вызовет более или менее дискретную реакцию системы по мере преодоления возмущающим фактором порога устойчивости [1193, 1194, 1196]. Краткие неустойчивые фазы эманации ассоциируются с переходными формами. Пунктуализм не противоречит постепенности филогенеза, настаивая лишь на значительной и закономерной неравномерности темпа эволюции. Это правило постулирует скоротечность и неустойчивость переходных форм и широкое распространение в филогенезе фаз относительной устойчивости архетипа таксона и направления его специализации.
Не всякая сальтация несет большой заряд новизны с ее разрушительными последствиями. Довольно резкие филогенетические перестройки могут быть связаны с неотенией и фетализацией [889, 1178, 1181], с переносом признаков с одного пола на другой [1193]. В этих случаях новое филогенетическое состояние имеет солидную исторически подготовленную базу, что в случае полноты материала легко опознать морфологически.
Правило «перемежающегося равновесия» было отчасти предвосхищено правилом происхождения нового наследственного признака В. Е. Руженцева, теоретически обоснованного М. А. Шишкиным [1178, 1180, 1181]. Это правило гласит: всякое новообразование в момент своего появления неопределенно и лишь пройдя испытание отбором, оно либо исчезает, либо становится полезным; изменчивость, отрицая наследственность, только при участии отбора сама становится наследственностью [887]. Формулировка В. Е. Руженцева указывает на исходную неустойчивость-нового признака (а значит, до известной степени и его обладателя) и связь устойчивости с адаптацией. На значение и приоритет правила В. Е. Руженцева недавно указал Л. Ван Вален [2424].
Правило дестабилизации (эманации). Это правило сформулировал В. В. Жерихин [314], базируясь прежде всего на экспериментах Г. X. Шапошникова [1147—1150, 2282] по детальному анализу смены адаптивных норм. Понятие «адаптивная норма» ввел И. И. Шмальгаузен [1184, 1192]. Его можно определить как ансамбль жизненных циклов, онтогенезов или их стадий, возникающих как исторически оправданный, устойчивый (и в этом смысле—адаптивный и наследственный) ответ на воздействия исторически типичной внутренней и внешней среды [514].
В опытах Г. X. Шапошникова по эволюции тлей в условиях смены кормового растения наблюдалась резкая дестабилизация многих морфологических признаков на фоне резко возросшего-уровня элиминации, достигавшего в отдельные моменты опыта 80—90 % численности популяции. За фазой дестабилизации последовала фаза стабилизации новой адаптивной нормы, сопровождавшаяся адаптацией к новому кормовому растению и понижением уровня элиминации до уровня контроля. Новая популяция имела морфологическую характеристику хорошего вида w практически полную репродуктивную изоляцию от контроля, т. е. предковой популяции. Смена адаптивной нормы, схематически изображенная на рис. 11, заняла 15 поколений тлей. Некоторые из результатов опыта недавно воспроизвел В. Б. Сапунов [912]; проведено еще несколько аналогичных экспериментов (см. 7.4).
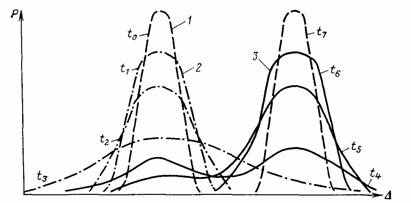
Рис. 11. Схема смены адаптивных норм в остром опыте Г. X. Шапошникова. Составил А. С. Раутиан. 1 — старая и новая устойчивые адаптивные нормы; 2 — стадии эманации (дестабилизации) старой адаптивной нормы; 3 — стадии стабилизации (специализации) новой адаптивной нормы; t0—t7 — последовательные моменты времени; по абсциссе отложена величина (количественный показатель) признака (Δ); по ординате—частота соответствующих количественных значений величины признака (Р)
Смена адаптивной нормы оказалась дискретной для популяции (частоты признаков) и градуальной для морфологических. рядов переходных форм, преемственно связавших старую и новую нормы. Плавное смещение популяционной кривой изменчивости в область исторически нетипичных значений признаков (в духе классической схемы движущего отбора—рис. 12, а) должно вызвать дестабилизацию популяции. Это и наблюдалось в опытах Д. К. Беляева [69] и сотрудников по искусственному движущему отбору (рис. 12, б). Градуальная смена адаптивной нормы популяции противоречит исторической обусловленности устойчивости ее адаптивной нормы. Снижение эффективности стабилизирующего отбора прежней нормы привело к ее дестабилизации. Снятие популяционных и некоторых организменных ограничений прежней нормы привело к мобилизации резерва изменчивости [1192], т. е. к повышению свободы выбора. Возросшая свобода творчества в сочетании с жестким стабилизирующим отбором устойчивых в новых условиях онтогенезов формируют новую адаптивную норму.
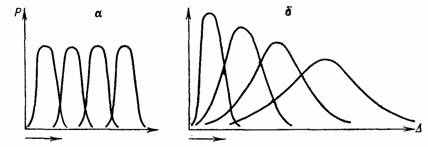
Рис. 12. Две версии движущей формы отбора: а — классическая схема (по Н. В. Тимофееву-Ресовскому, Н. Н. Воронцову и А. В. Я6локову); б — схема движущей формы искусственного отбора в острых опытах Д. К. Беляева и сотрудников: наблюдается дестабилизирующий эффект движущей формы отбора (составил А. С. Раутиан по данным Д, К. Беляева). Стрелкой показано направление вектора отбора и времени
Эффективность движущего отбора в дестабилизированной популяции должна быть исчезающе мала:
1) дестабилизация понижает наследуемость признаков и эффективность отбора по ним;
2) эффективный отбор признаков с низкой наследуемостью требует длительного времени [577], а затяжная дестабилизация— верная гибель;
3) чем сильнее движущий отбор смещает популяцию за пределы адаптивной нормы, тем слабее его направляющая роль. Область поисков новой адаптивной нормы ограничивается структурой обратимо мобилизуемого резерва изменчивости, определяющейся, в свою очередь, структурой прежней нормы, а не эффективностью движущего отбора.
Палеонтология не осталась в стороне от исследования дестабилизации, особенно трудного для нее по причине редкости переходных форм. Одно из филогенетических правил Э. Д. Копа [1457] гласит: перед моментом вымирания группа, как правило, повышает свое таксономическое разнообразие. Речь идет, очевидно, о неоднократных попытках вымирающей группы выйти из экологического кризиса путем смены направления специализаций (немыслимого без дестабилизации). Но все попытки кончались лишь формированием короткоживущих таксонов. Это правило-связывает процессы дестабилизации как с вымиранием, так и с типогенезом. Правило возникновения нового наследственного признака В. Е. Руженцева, концепции квантовой эволюции [952,. 1150, 2282] и пунктуализма явились важными шагами в понимании чередования фаз специализации и эманации, а также неустойчивости и скоротечности последней. Наконец, само правило дестабилизации сформулировал палеонтолог В. В. Жерихин. На ископаемом материале продемонстрированы дестабилизации целых экосистем и связанные с ними волны вымирания и появления новых групп [367, 368, 457].
Правило прогрессивной специализации. Если в процессе развития устойчивость и защищенность системы от внешних и внутренних возмущений растет, т. е. идет процесс прогрессивного приспособления в избранном направлении, то мы встречаемся со специализацией [313]. Говорить о постоянстве направления специализации можно только применительно к избранному уровню-рассмотрения, поскольку всякое изменение состояния эволюирую-щей системы требует известного нарушения устойчивости — дестабилизации, а следовательно, и нарушения направленности. Иерархия корреляционных систем организма и их координированных изменений в филогенезе [1192, 1196, 1197]—та биологическая реальность, которая стоит за этими уровнями рассмотрения, отраженными, в частности, в таксономической иерархии. Низшим уровнем специализации будет процесс стабилизации фенотипов определенного класса.
Прогрессивная, длительное время не прерывающаяся на данном уровне рассмотрения, специализация, сохраняющая архетип таксона соответствующего ранга и направление адаптивного совершенствования, является воплощением постепенного преемственного и наиболее безопасного в отношении вымирания развития, составляющего основной фон филогенетических событий. Напротив, резкое возмущение системы, крупное приращение структурной новизны ослабляет преемственность, форсирует забвение прежнего адаптивного опыта, нарушает устойчивость канала развития— филокреода [647], меняя направление специализации.
Особенно ярко прогрессивная специализация проявляется в явлениях параллелизма, когда независимые потомки общего предка сохраняют тенденцию приобретения гомологичных признаков и целых функциональных аппаратов. Эта тенденция нашла отражение в явлениях гомоплазии (Дж. Рей Ланкастер), понятиях латентной, потенциальной (Г. Осборн), косвенной или параллельной (В. Е. Руженцев) гомологии [291, 887]. Об этом же говорят правила гомологических и гетерологических рядов Э. Копа [1457], .схематически показанных на рис. 13—15. Явления параллельной и итеративной эволюции, в частности правило параллельного достижения разными родственными филетическими линиями приблизительно одного уровня организации [1033, 1457, 2287], нашли отражение в представлении о вертикальной и горизонтальной классификации [883, 1033]. Эти эмпирические обобщения свидетельствуют о большой роли начальных условий развития и преемственности, ведущих к проявлению одних и тех же латентных потенций в параллельных линиях специализации [74, 592, 975]. Вероятный механизм действия начальных условий был рассмотрен Н. Н. Иорданским [444, 445] и будет описан при характеристике правила ключевого ароморфоза.
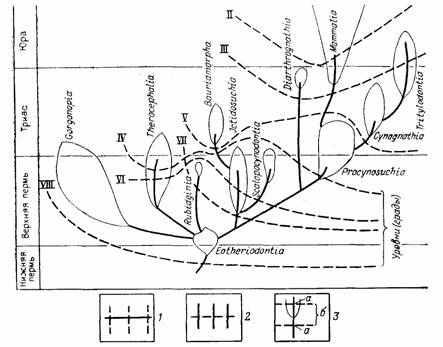
Рис. 13. Аристогенез в эволюции териодонтов. По Л. П. Татаринову. Кривые, пересекающие филстические линии, соответствуют уровням (градам), на которых приобретались характерные для млекопитающих признаки; II — звукопроводящий аппарат из трех слуховых косточек, III — челюстное сочленение между зубной и чешуйчатой костями, IV — мягкие, снабженные собственной мускулатурой губы, V — несколько расширенные большие полушария головного мозга, VI — зона усиленного кровоснабжения на губах, связанная, по-видимому, с формированием зачаточных вибрисс, VII — трехбугорчатые щечные зубы, VIII—верхние обонятельные раковины; ряды Э. Д. Копа; 1—гомологические, 2 — гетерологические; 3 — сопряжения филетических линий с уровнями (градами): а—мутации В. Ваагена, ректиградации или аристогены Г. Ф. Осборна, б — аллойометроны Г. Ф. Осборна