
Лекции деление клетки (митоз, мейоз и их отличиях) и индивидуальное развитие (образовании гамет и оплодотворения и краткий обзор эмбрионального развития)
| Вид материала | Лекции |
СодержаниеКраткий обзор этапов гаметогенеза Caenorhabditis elegans Гомеозисные гены |
- Лекции деление клетки (митоз, мейоз и их отличиях) и индивидуальное развитие (образовании, 155.65kb.
- Лекция №3 Механизмы клеточного деления. Митотический цикл и его регуляция. Митоз. Мейоз, 199.23kb.
- Уроков с использованием икт урок по теме «Деление клетки. Митоз», 40.88kb.
- Тема : Деление клетки основа размножения, роста и развития организмов, 105.53kb.
- Митоз (непрямое деление) является самым распространенным способом деления клеток., 29.36kb.
- Способы деления эукариотических клеток: митоз, мейоз, амитоз, 51.33kb.
- Цель: изучить закономерности эмбрионального развития у различных позво, 128.84kb.
- Митоз. Фазы митоза. Значение митоза. Цитокинез растений и животных. Другие виды нередукционного, 117.9kb.
- Лекция 15. Мейоз, гаметогенез Мейоз, 145.8kb.
- План Расширения понятия угла и дуги, их измерение. Тригонометрические функции в Индии., 98.28kb.
| Митоз, мейоз, гаметогенез, оплодотворение и эмбриональное развитие
| |
| Тема лекции – деление клетки (митоз, мейоз и их отличиях) и индивидуальное развитие (образовании гамет и оплодотворения и краткий обзор эмбрионального развития). Клетка в своей жизни проходит разные состояния: фазу роста и фазы подготовки к делению и деления. Клеточный цикл – переход от деления к синтезу веществ, составляющих клетку, а затем опять к делению – можно представить на схеме в виде цикла, в котором выделяют несколько фаз.  После деления клетка вступает в фазу синтеза белков и роста, эту фазу называют G1. Часть клеток из этой фазы переходит в фазу G0, эти клетки функционируют и потом погибают без деления (например, эритроциты). Но большинство клеток, накопив необходимые вещества и восстановив свой размер, а иногда и без изменения размеров после предыдущего деления, начинают подготовку к следующему делению. Эта фаза называется фаза S – фаза синтеза ДНК, затем, когда хромосомы удвоились, клетка переходит в фазу G2 – фазу подготовки в митозу. Затем происходит митоз (деление клетки), и цикл повторяется заново. Фазы G1, G2, S вместе называются интерфазой (т.е. фазой между делениями клетки).  Жизнь клетки и переход от одной фазы клеточного цикла к другой регулируется изменением концентраций белков циклинов , как это показано на рисунке. 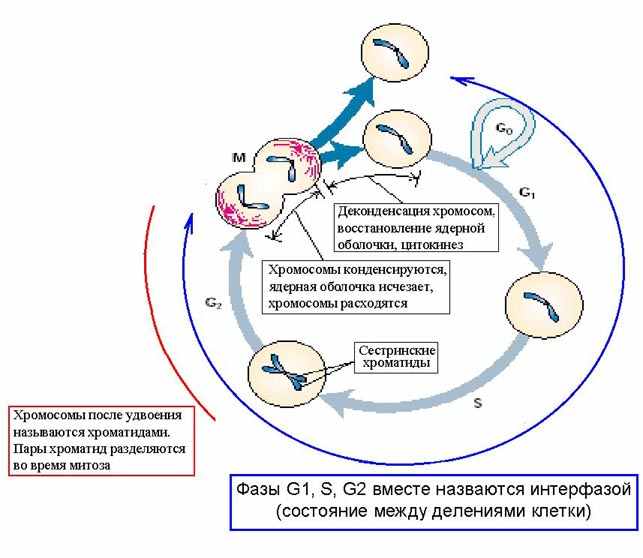 При подготовке к делению происходит репликация ДНК, на каждой хромосоме синтезируется ее копия. Пока эти хромосомы после удвоения не расходятся, каждая хромосома в этой паре называется хроматидой. После репликации ДНК конденсируется, хромосомы приобретают более компактную укладку, и в таком состоянии их можно увидеть в световом микроскопе. Между делениями эти хромосомы не столь конденсированы и в большей степени расплетены. Понятно, что в конденсированном состоянии им трудно функционировать. Хромосома имеет вид в виде буквы Х только во время одной из стадий митоза. Раньше считалось, что между делениями клетки хромосомная ДНК ( хроматин ) находится в полностью расплетенном состоянии, но сейчас выясняется, что структура хромосом достаточно сложная и степень деконденсации хроматина между делениями не очень велика. 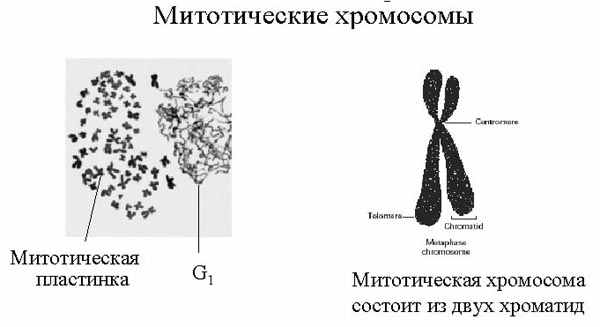 Процесс деления, при котором исходно диплоидная клетка дает две дочерние, также диплоидные, клетки, называется митозом. Имеющиеся в клетке хромосомы удваиваются, выстраиваются в клетке, образуя митотическую пластинку, к ним прикреплены нити веретена деления, которые растягиваются к полюсам клетки и клетка делится, образуя две копии исходного набора.  При образовании гамет, т.е. половых клеток – сперматозоидов и яйцеклеток – происходит деление клетки, называемое мейозом. Исходная клетка имеет диплоидный набор хромосом, которые затем удваиваются. Но, если при митозе в каждой хромосоме хроматиды просто расходятся, то при мейозе хромосома (состоящая из двух хроматид) тесно переплетается своими частями с другой, гомологичной ей хромосомой (также состоящей из двух хроматид), и происходит кроссинговер - обмен гомологичными участками хромосом. Затем уже новые хромосомы с перемешанными «мамиными» и «папиными» генами расходятся и образуются клетки с диплоидным набором хромосом, но состав этих хромосом уже отличается от исходного, в них произошла рекомбинация . Завершается первое деление мейоза, и второе деление мейоза происходит без синтеза ДНК, поэтому при этом делении количество ДНК уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки образуются четыре гаплоидных клетки. Фазы деления клетки, которые следуют за интерфазой, называются профаза, метафаза, анафаза, телофаза и после деления опять интерфаза. 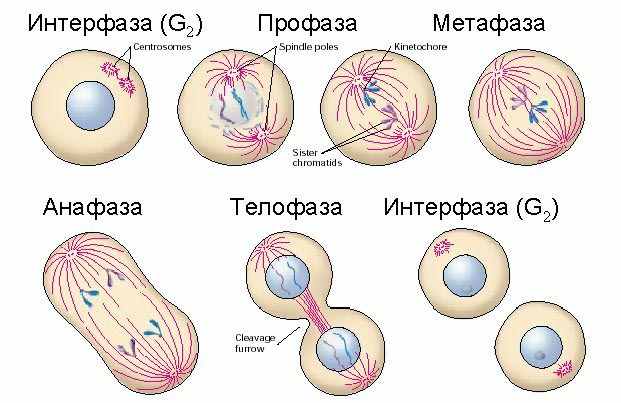 При мейозе фазы называется также, но указывается к какому делению мейоза она относится. Кроссинговер – обмен частями между гомологичными хромосомами – происходит в профазе первого деления мейоза (профаза I), которая включает следующие этапы: лептотена, зиготена, пахитена, диплотена, диакинез. Процессы, происходящие при этом в клетке, подробно описаны в учебнике Макеева, и их следует знать. 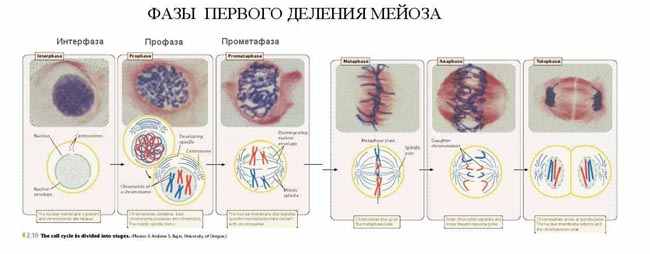 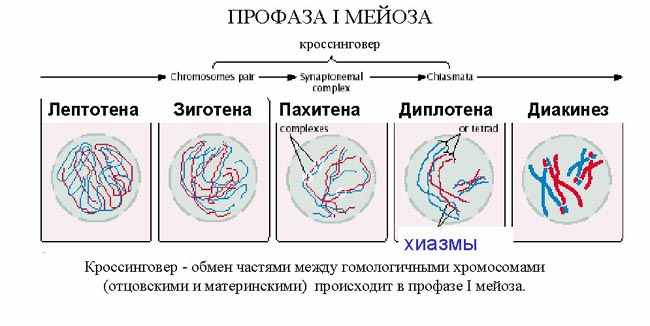 Краткий обзор этапов гаметогенеза Гаметогенез подразделяется на сперматогенез (процесс образования сперматозоидов у самцов) и оогенез (процесс образования яйцеклетки). По тому, что происходит с ДНК, эти процессы практически не отличаются: одна исходная диплоидная клетка дает четыре гаплоидные. Однако, по тому, что происходит с цитоплазмой, эти процессы кардинально различаются. В яйцеклетке накапливаются питательные вещества, необходимые в дальнейшем для развития зародыша, поэтому яйцеклетка – это очень крупная клетка, и когда она делится, цель – сохранить питательные вещества для будущего зародыша, поэтому деление цитоплазмы несимметрично. Для того чтобы сохранить все запасы цитоплазмы и при этом избавиться от ненужного генетического материала, от цитоплазмы отделяются полярные тельца, которые содержат очень мало цитоплазмы, но позволяют поделить хромосомный набор. Полярные тельца отделяются при первом и втором делении мейоза (подробнее о том, что происходит с полярными тельцами растений – в Макееве)  Исходная клетка, из которой в последствии образуется зрелая яйцеклетка, называется ооцитом первого порядка. После деления из него образуется ооцит второго порядка и первое полярное тельце. Затем происходит второе деление мейоза, в результате образуется гаплоидный оотид и второе полярное тельце. Первое полярное тельце за это время тоже успевает поделиться, таким образом всего получается три гаплоидных полярных тельца. В оотиде происходят некоторые процессы созревания и он превращается в яйцеклетку. Она содержащая почти всю цитоплазму исходного ооцита, но гаплоидный набор хромосом. Эти хромосомы уже прошли рекомбинацию, т.е. если исходно клетки содержат одну хромосому от мамы, одну от папы, то в зрелой яйцеклетке в каждой хромосоме чередуются куски, полученные от одного и второго родителя. При сперматогенезе цитоплазма исходного сперматоцита первого порядка делится (первое деление мейоза) поровну между клетками, давая сперматоциты второго порядка. Второе деление мейоза приводит к образованию гаплоидных сперматоцитов второго порядка. Затем происходит созревание без деления клетки, большая часть цитоплазмы отбрасывается, и получаются сперматозоиды, содержащие гаплоидный набор хромосом очень мало цитоплазмы. Ниже представлена фотография сперматозоида человека и схема его строения. Сперматозоиды животных имеют одинаковое принципиальное строение, но могут отличаться формой и размером. Сперматозоид имеет головку, в которую плотно упакована ДНК. Головка сперматозоида окружена очень тонким слоем цитоплазмы. На ее переднем конце находится структура, называемая акросомой. Эта структура содержит ферменты, позволяющие сперматозоиду проникнуть через оболочку яйцеклетки. Сперматозоид имеет хвостик. Часть хвостика, прилегающая к головке ("шейка"), окружена митохондриями. Они необходимы, чтобы обеспечить биение хвостика и движение сперматозоида в желательном ему направлении. На сперматозоиде имеется для выбора направления движения хеморецепторы, сходные с обонятельными клетками. 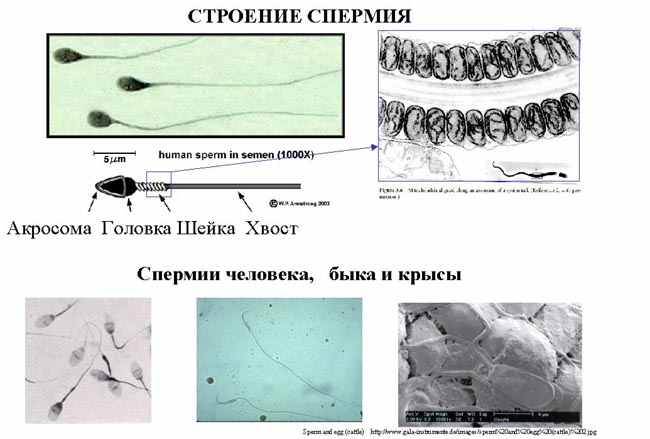 Созревание спермиев происходит в семенных канальцах тестикул. При превращении исходной клетки, сперматогония, в сперматоцит, сперматиды и зрелый сперматозоид происходит перемещение клетки от базальной мембраны семенного канатика к его полости. После созревания сперматозоиды отделяются, попадая в просвет семенных канальцев, и готовы к движению в поисках яйцеклетки и оплодотворению. Процесс созревания длится примерно три месяца. У млекопитающих у особей мужского пола процесс созревания сперматозоидов – сперматогенез – начинается с возраста половой зрелости и продолжается затем до глубокой старости. 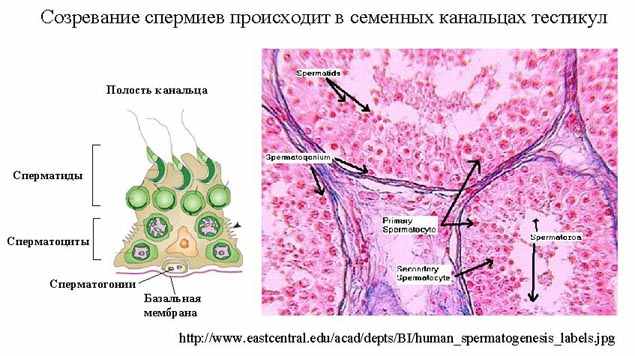 Существенно отличается процесс созревания яйцеклетки – оогенез. Во время эмбрионального развития млекопитающих возникает большое количество яйцеклеток, и к рождению самки в ее яичниках уже находится порядка 200-300 тысяч яйцеклеток, остановившихся на первой стадии деления мейоза. В период полового созревания яйцеклетки начинают реагировать на половые гормоны, Регулярные циклические изменения гормонов впоследствии вызывают созревание яйцеклетки, обычно одной, иногда двух или больше. Когда для лечения бесплодия женщине делают инъекции половых гормонов, чтобы индуцировать созревание яйцеклеток, избыток этих гормонов может привести к созреванию нескольких яйцеклеток, и как следствие этого – многоплодной беременности. Яйцеклетка созревает в пузырьке, называемом фолликулом.  За всю жизнь у женщин современных индустриализованных стран созревает всего 400-500 яйцеклеток, у женщин традиционной культуры – в племенах охотников-собирателей – менее 200 штук. Это связано с различиями в традиции деторождения: у европейских женщин рождается в среднем 1-2 ребенка, которых она кормит в среднем 3 -5 месяцев, (а известно, что лактация тормозит восстановление месячных циклов после родов), то есть у нее больший период времени остается для созревания яйцеклеток и прохождения менструальных циклов; в это же самое время у бушменов женщины рожают в среднем по 5 детей, они не делают абортов, в отличие от западных женщин, и они кормят грудью по 3-4 года, при этом овуляция тормозится, поэтому месячных циклов у них в 2 раза меньше, чем у западных женщин. Большее количество овуляторных циклов ведет к повышению риска заболевания репродуктивных органов у женщин, так как каждая овуляция связана с делением клеток, а чем больше делений – тем больше может возникнуть мутаций, ведущих к появлению злокачественных образований. Месячные циклы у женщины регулируются изменением концентрации гормонов (верхний график на рисунке). Под действием гормонов один из покоящихся фолликулов (пузырьков) с яйцеклеткой начинает развиваться. Через несколько дней фолликул лопается и из него выходит зрелая яйцеклетка. Этот процесс называется овуляцией. Слизистая оболочка матки (эндометрий) при этом разрастается, готовясь принять оплодотворенную яйцеклетку. Если беременность не наступает, происходит дегенерация и отторжение верхнего слоя эндометрия, сопровождающееся кровотечением. Во время овуляции у женщины происходит повышение так называемой базальной температуры (то есть температуры, измеряемой ректально и вагинально сразу после пробуждения) на несколько десятых градуса (нижний график на рисунке), потом она может упасть или остаться слегка повышенной до начала менструации. У каждой женщины колебания базальной температуры индивидуальны, но более или менее постоянны при установившемся месячном цикле. Таким образом по изменению температуры можно примерно судить, когда происходит овуляция. 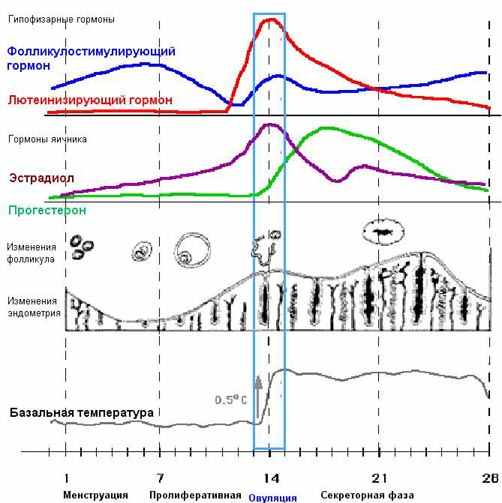 Ошибки при определении сроков овуляции по базальной температуре могут возникать из-за не связанных с месячным циклом изменений температуры (например, при гриппе или другом заболевании, дающем подъем температуры) или из-за сбоев цикла, которые могут возникнуть у женщины при перемене климата, стрессе или под влиянием других факторов. Пример изменения температуры в одном месячном цикле представлен на рисунке: 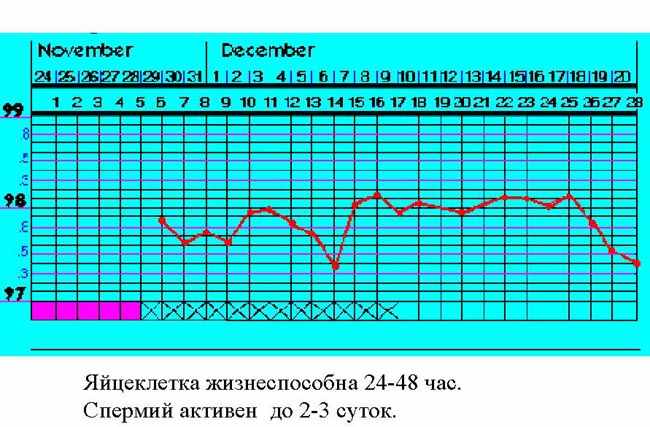 Яйцеклетка после выхода из фолликула сохраняет жизнеспособность примерно 24-48 часов. Спермии же после попадания в половые пути женщины жизнеспособны до 2-3 суток, далее они могут быть подвижны, но не способны к оплодотворению. Поэтому оплодотворение возможно в течение 2-3 дней до и 1-2 дней после овуляции. В остальное время зачатие произойти не может. Но на самом деле скачок температуры происходит не точно при овуляции, а при изменении концентрации гормонов, овуляцию вызывающих, поэтому точность определения дня овуляции по температурному графику составляет примерно 2 дня. Поэтому оплодотворение может произойти в 3+2=5 дней до овуляции и 2+2=4 дня после овуляции дней цикла. Осторожные люди прибавляют еще по 1-2 дня с каждой стороны. Остальные дни считаются "безопасными". Хотелось бы отметить, что цикл подчиняется эмоциональной регуляции, например, во время войны из-за тяжелой жизни, недоедания у женщин прекращались менструации, это явление называется "аменорея военного времени". Однако описаны случаи, когда муж приезжал домой с фронта на 2 дня, за эти 2 дня у женщины происходила овуляция независимо от фазы цикла, и впоследствии рождался ребенок. О том, что физиологические процессы достаточно сильно могут регулироваться нервной системой, показывает процесс родов у обезьян. У человека первые роды длятся примерно 24 часа, а у обезьян всего несколько часов, причем начинаются они обычно во время, когда стадо находится на стоянке. То есть к утру, когда стадо собирается отправляться в путь, мама готова путешествовать дальше с новорожденным. Если по каким-то причинам процесс родов к утру не завершился, а стадо уже готово идти дальше, то роды останавливаются, так как стадные животные не должны отставать от своих сородичей, и уже потом при новой остановке, роды возобновляются. Процесс проникновения сперматозоидов в яйцеклетку называется оплодотворением. Яйцеклетка окружена несколькими оболочками, структура которых такова, что только сперматозоид собственного вида может попасть в яйцеклетку. После оплодотворения оболочки яйцеклетки меняются и другие сперматозоиды уже не могут в нее проникнуть. 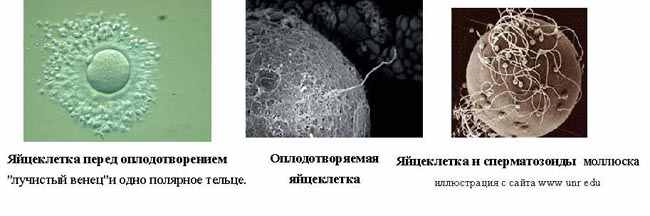 У некоторых видов внутрь яйцеклетки могут проникнуть несколько сперматозоидов, но все равно в слиянии ядер участвует только один из них. При оплодотворении в яйцеклетку проникает только ядро сперматозоида, хвостик же вместе митохондриями отбрасывается, и в клетку не попадает. Поэтому митохондриальную ДНК все животные наследуют только от матери. Оплодотворенное яйцо называют зиготой (от греч. зиготос – соединенный вместе). После оплодотворения происходит деление клетки, восстановившей диплоидный набор хромосом. первое и несколько последующих делений яйцеклетки происходят без увеличения размера клеток, поэтому процесс называется дроблением яйцеклетки. Эмбрион (греч. "зародыш") - ранняя стадия развития живого организма от начала дробления яйцеклетки до выхода из яйца или из материнского организма (в акушерстве, в отличие от эмбриологии, термин эмбрион используют только для первых 8 недель развития, после 8-й недели называют плодом).  Эмбриогенез (эмбриональное развитие) является частью онтогенеза (индивидуального развития) – развития организма от образования зиготы до его смерти. Эмбриогенез - это процесс, в котором презумптивные зачатки занимают свои дефинитивные места. Из школьного вы помните, что при развитии эмбриона ланцетника образуется бластула (полый клеточный шарик), из которой получается двуслойная гаструла путем инвагинации (впячивания) одной стороны бластулы внутрь. 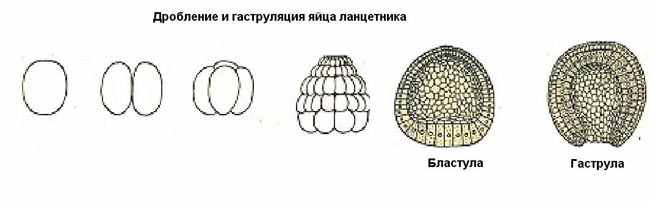 У млекопитающих процесс происходит несколько иным образом. Дробление яйцеклетки у них приводит к образованию комочка клеток, называемого морулой. Морула подразделяется на внутреннюю часть, из которой потом развивается сам зародыш, и наружную часть, образующую полый пузырек, называемый трофобластом. Дальнейшее развитие приводит к формированию трехслойного зародыша, состоящего из внутреннего слоя – энтодермы, внешнего слоя – эктодермы, и третьего слоя между ними – мезодермы. Из каждого слоя впоследствии образуются определенные ткани и органы. На фотографии ниже изображен червь нематода Caenorhabditis elegans (произносится как "ценорабдитис элеганс"), ближайший родственник аскариды, который интересен тем, что имеет строго фиксированное число клеток. Это дает возможность установить происхождение каждой из клеток, какая клетка из какой получилась. На рисунке показана схема происхождения разных клеток в эмбриональном развитии C.elegans. 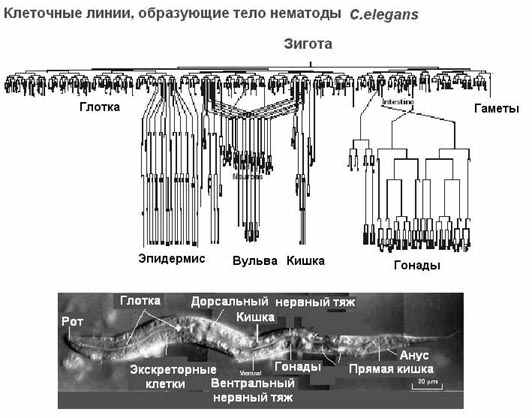 Во время развития зародыша происходит дифференциация и перемещение его клеток с образованием тканей и органов. Рассмотрим на примере мухи-дрозофилы этот процесс. В развитии мухи происходит последовательная смена форм, значительно отличающихся друг от друга: яйцо, личинка, куколка и имаго (взрослая особь). Такое развитие называется развитием с метаморфозом (метаморфоз – греч. "изменение формы").  Цитоплазма яйцеклетки не гомогенна, в ней асимметрично распределены различные биологически активные компоненты. У эмбриона уже определены оси тела, число и ориентация сегментов тела, из которых затем развиваются части тела взрослой мухи. Эти процесс контролируются различными наборами генов, которые называются морфогены. Они кодируют белки, которые регулируют экспрессию других генов, отвечающих за формирование органов. Градиент концентрации белков-морфогенов определяет передне-заднюю и дорсо-вентральную (спино-брюшную) оси тела. У дрозофилы в определении дорсо-вентральной оси тела участвуют 12, главный из них ген dorsal. Белок Dorsal сконцентрирован в цитоплазме на той стороне эмбриона, которая станет спинной, и проникает в ядро клеток на брюшной стороне, активируя группы генов, продукты которых необходимы для определения осей тела. Детерминация передне-задней оси тела контролируется другими генами. Один из них называется bicoid, его мРНК сконцентрирована в цитоплазме передней части яйца (фиксирована своим 3'-концом). В результате при трансляции возникает градиент концентрации белка Bicoid от переднего к заднему концу яйца. Градиент поддерживается продолжительным периодом синтеза белка и его коротким временем жизни. Bicoid, так же как и Dorsal, морфоген, он активирует гены, которые необходимы для развития головы и грудных структур. Его экспрессия ингибируется продуктом гена nanos, сконцентрированного на заднем конце эмбриона. На следующем этапе включаются гены сегментации. Они контролируют дифференциацию эмбриона на индивидуальные сегменты. После оплодотворения транскрибируется около 25 генов сегментации, их экспрессия регулируется градиентами белков Bicoid и Nanos. 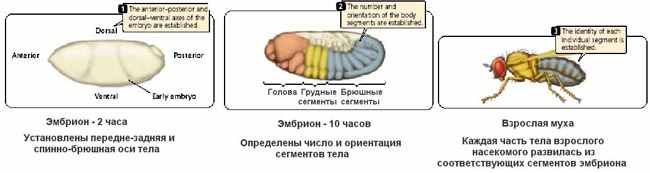 Гомеозисные гены После сегментации и установления ориентации сегментов активируются так называемые геомеозисные гены. Различные их наборы активируются специфическими соотношениями концентраций белков, упоминавшихся выше. Продукты гомеозисных генов активируют другие гены, которые определяют сегмент-специфичные особенности. Глаза в норме возникают только на головном сегменте, а ноги – только на грудных сегментах. Гомеозисные гены кодируют регуляторные белки, связывающиеся с ДНК. Каждый из них содержит кластер нуклеотидов, называемый гомеобоксом, которые сходен во всех гомеотических генах. Он содержит 180 нуклеотидов и кодирует 60 аминокислот, функционирующих как ДНК-связывающий домен. У дрозофилы имеется два основных кластера гомеотических генов: комплекс Antennapedia (5 генов у дрозофилы) который определяет развитие головы и передних торакальных сегментов, и комплекс Bithorax (3 гена) который контролирует развитие задних торакальных и брюшных сегментов. Порядок расположения генов тот же, что и сегментов, в которых они экспрессируются. Впервые мутации гомеозисных генов были идентифицированы в 1894 году, когда Уильям Бэтсон заметил, что у растений иногда части цветка появляются на неправильных местах. Гомеозисные гены как бы определяют адрес клетки в конкретном сегменте, сообщая клеткам, в каком районе они находятся. Когда они мутируют, клетки получают "ложный адрес" и ведут себя так, будто они находятся в другом месте эмбриона Нарушения в работе гомеозисных генов (вызванные мутациями или внешними воздействиями) нарушают формирование структур тела и могут привести, например, к образованию глаз на лапках у мухи, или к тому, что вместо антенн на голову у нее вырастут ноги (как это показано на рисунке). У человека найдены мутации в гомеозисных генах, приводящие к недоразвитию зубов, например, и к другим, более тяжелым нарушениям. 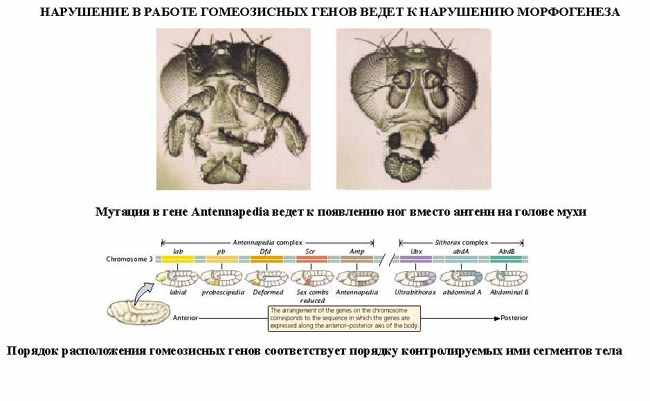 После того, как были открыты и изучены геомео-гены дрозофилы, сходные гены были найдены у всех других животных от нематоды до человека. У млекопитающих они называются Hox генами (гомеобокс-содержащими генами) и также кодируют белки, регулирующие транскрипции и определяющие структуры тела и их положение в передне-заднем направлении. 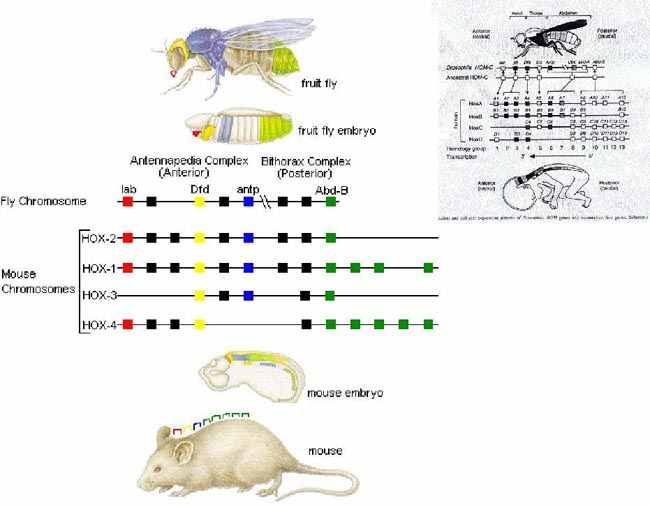 Таким образом, в эмбриональном развитии исходный градиент белков и мРНК в яйцеклетке стимулирует локальную экспрессию генов эмбриона, которая ведет дальнейшей дифференциации генной экспрессии и определяет судьбу клеток развивающегося эмбриона. Процесс, в котором формируются конечности, ткани и органы, называется морфогенезом, и определяется последовательностью переключения экспрессии групп генов, однако эти гены пока не столь детально изучены. В процессе эмбриогенеза осуществление записанной в генах программы развития происходит в конкретных условиях среды. Взаимодействие генов и среды можно описать на следующей модели. Эмбриональное развитие можно сравнить с шариком, катящимся по наклонной поверхности с разными желобками. Такое представление эмбрионального развития, названное эпигенетическим ландшафтом, был предложен Кондратом Уоддингтоном. 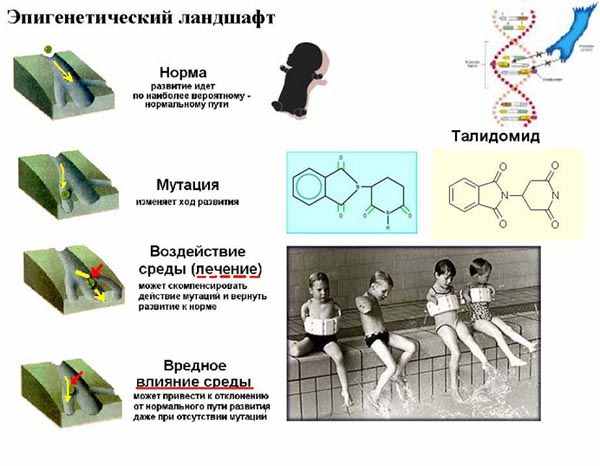 Самый глубокий желобок (соответствующий наиболее вероятному пути) определяет нормальное развитие организма. Но у основного желобка есть много разветвлений, менее глубоких, соответствующих патологии, аномальному развитию организма, по ним шарик покатится с меньшей долей вероятности. Мутации меняют соотношение вероятностей разных путей (на рисунке – меняется глубина желобков), и увеличивают вероятность развития по "неправильному" патологическому пути. Однако в части случаев воздействие среды (лечение) может скомпенсировать дефект и вернуть организм на нормальный путь развития. Например, фенилкетонурия – наследственная болезнь, которую можно лечить. Суть болезни заключается в том, что у больных отсутствует фермент фенилаланингидроксилаза, превращающий аминокислоту фенилаланин в другую аминокислоту, тирозин. При блокировании нормальных путей катаболизма фенилаланина его превращение идет другими путями, обычно играющими второстепенную роль. Фенилаланин превращается в кетокислоту фенилпируват (фенилпировиноградная кислота) и другие продукты. Избыточные количества фенилпирувата легко определить по анализу мочи, и такой анализ проводится всем новорожденным в нашей стране. Одним из симптомов этой болезни является развитие умственной отсталости, которое во взрослом состоянии уже необратимо. Лечить болезнь можно в детстве специальной диетой, при которой в организме не из чего будет вырабатывать пировиноградную кислоту. Частота заболевания около 1:10 000 новорожденных, и чем раньше начато лечение – тем лучше результаты. Именно поэтому проводится тотальная диагностика новорожденных. Интересно то, что если ребенок перестает придерживаться лечебной диеты, то болезнь опять станет прогрессировать. Поэтому диету надо соблюдать до остановки физиологического роста, примерно до 20 лет, когда токсичное воздействие будет менее опасным. Больные фенилкетонурией при беременности обязательно должны соблюдать диету, так как иначе плод будет отравлен из-за нарушения обмена веществ у матери. Таким образом, при лечении, то есть полезном воздействии внешней среды, можно вернуть развитие организма в нормальное русло. Но действие окружающей среды может быть и вредным, то есть у организма под действием внешней среды возникают отклонения развития при совершенно нормальных генах. Для примера рассмотрим один случай. В 60-х годах в Германии было сильно разрекламировано новое снотворное под названием талидомид. Среди принимавших новое лекарство были беременные женщины. Спустя некоторое время было замечено, что в стране стало рождаться много детей с патологией конечностей. У них отсутствовали длинные кости конечностей, то есть прямо от основания тела начинались кисти или ступни. Раньше такое заболевание встречалась один раз на несколько тысяч новорожденных, и вдруг такой всплеск. Начали проводиться исследования, и выяснилось, что причина в новом лекарстве. Как оказалось, талидомид имеет большое сродство к гуанину. Взаимодействуя с ДНК, он может приводить к функциональным нарушениям. Промотор гена, отвечающего за рост и развитие длинных конечностей, содержит большое количество гуанина, таким образом, талидомид нарушает работу этих генов, и зачатки костей длинных конечностей так и не начинают развиваться. Многие из этих детей не выжили, часть из тех, кто выжил, ведут жизнь инвалидов, но есть среди них люди, которые, несмотря на инвалидность, реализовали свои возможности. После талидомидной трагедии все новые лекарства проверяют на тератогенную (вызывающую нарушения развития плода) активность, и для каждого препарата указано, можно ли его принимать беременным. Однако следует учитывать, что во время беременности, особенно на ранних этапах, женщина не должна принимать лекарства, не посоветовавшись с врачом, из-за возможных вредных воздействий на плод. В настоящее время уровень тяжелых врожденных уродств составляет 1-2%, из них около трети по генетическим причинам, около трети – из-за воздействий среды, и для трети причина неизвестна. Подбирая условия среды, соответствующие индивидуальным особенностям организма, можно скомпенсировать часть врожденных дефектов. Взаимодействие генов и среды в процессе индивидуального развития можно сравнить с игрой в карты: хороший игрок может выиграть и с плохими картами. |
