
Флора и фитогеография печеночников (marchantiophyta, anthocerotophyta) камчатки и прилегающих островов 03. 00. 05 ботаника
| Вид материала | Автореферат диссертации |
- Флора и растительность копаней ярославской области 03. 00. 05 ботаника, 463.62kb.
- Сохранение биоразнообразия камчатки и прилегающих морей южно-Камчатский заказник: изменения, 45.7kb.
- Гидрофильная флора сосудистых растений как модельный объект для инвентаризации и анализа, 555.67kb.
- На семинаре предполагается рассмотреть следующие основные темы, 47.38kb.
- Кузьмицкого Алексея Алексеевича, формирование по сути однопартийного представительного, 1278.31kb.
- И с Россией Андреевский флаг, 457.61kb.
- Соломон Шульман, 2770.57kb.
- Географическое положение, 122.67kb.
- Дальнего Востока России. Официальный язык конференции русский доклад, 45.87kb.
- Флора Вафина (творческий псевдоним-Флора) автор слов и музыки, исполнитель романсов,, 133.03kb.
Благодаря свойству растительного покрова - мозаичности, любое выделение флористических районов отличается условностью, поскольку границы их весьма расплывчаты. Вместе с тем, очевидно, что не может существовать двух совершенно одинаковых локальных флор. Существуют достоверные факты, говорящие о различии во флоре группировок печеночников одних и тех же поясов (например, горной тундры) на юге и на севере п-ова или, тем более, островных массивов. Так, на мелкоземе вблизи снежников в тундрах на севере п-ова произрастает Schofieldia monticola, не известная южнее 58° с.ш., несмотря на наличие подходящих субстратов и сходных сообществ. Cryptocolepsis imbricata, произрастающий на Камчатке и Северных Курилах на вулканическом пепле и скалах, достигает лишь 56° с.ш., но не известен севернее. Porella platyphylla, найденная на о-ве Медный на камнях в истоках ручья в шикшевой тундре - местообитании более чем обычном в регионе - не обнаружена ни на других островах, ни на полуострове. Эти различия объясняются, главным образом, историей миграций печеночников, характером и степенью развития оледенений, вулканической деятельностью, случайным дальним переносом и, наконец, районными климатическими девиациями.
 |
| Рис. 8. Корреляция между географическими спектрами флор избранных районов, при нижней границе коэффициента 99%. |
В целом, как широтные, так и долготные спектры весьма однородны. В широтном отношении выделяется лишь Западная Камчатка, за счет снижения количества арктических видов и повышения арктобореальных и бореальных, что является следствием равнинного характера рельефа. В долготном отношении выделяется Южная Камчатка, где уменьшается доля циркумполярных видов за счет увеличения восточноазиатских и амфиокеанических. Отличие можно истолковать южным и океаническим положением этого района, но в таком случае приходится объяснять, почему сходные тенденции не наблюдаются на Северных Курилах и Командорах. Вероятно, причиной этого является их островное положение, когда флористический состав смещается в сторону преобладания активно размножающихся таксонов, имеющих (как следствие) широкий циркумполярный ареал.
Для того чтобы оценить географическую связь с ближайшими районами, в анализ, кроме флористических подразделений Камчатки, были вовлечены о-ва Кунашир и Итуруп, южная часть о-ва Сахалин, две естественные группы островов Алеутской дуги (Атту-Агатту и Амчитка-Унимак) и п-ов Аляска (Рис. 8).
При уровне сходства в 99% выделяется 3 связанные друг с другом клады. Причины, объединяющие каждую их них, различны и основываются на сходстве рельефа, климатических условий или положением внутри единого миграционного потока. Первая объединяет Северную, Центральную, Восточную Камчатку, Северные Курилы и Командоры. Это объединение, по-видимому, является следствием сходства рельефа при довольно жестком климате верхних поясов гор и выражается в наличии большого количества видов, тяготеющих к горам. Вторая клада объединяет Южную Камчатку, Курилы, Командоры и комплекс островов Амчитка-Унимак. Возможная причина объединения - сходный океанический, холодный, с жестким ветровым режимом климат и наличие видов приокеанического распространения. Третья охватывает Южную Камчатку, п-ов Аляска и комплексы островов Амчитка-Унимак и Атту-Агатту. Причина объединения - океанический влажный климат и положение на пути миграции видов из Азии в Америку. Следствие - наличие видов реликтового, более южного, распространения и приокеанических таксонов. Разумеется, приведенные здесь объяснения предположительны, но бесспорно существование отмеченных связей.
При нижнем пороге коэффициента корреляции в 97% происходит объединение всех районов, относящихся к Циркумбореальной области. Районы, занимающие пограничное положение - Кунашир и Итуруп - образуют отдельную кладу. При нижнем пороге в 95% флора о-ва Итуруп и Южного Сахалина связывается с типичными циркумбореальными флорами расположенными севернее. Наконец, при пороге корреляции в 70% соединяются все вовлеченные в анализ флоры. Таким образом, отчетливые связи, как на региональном, так и на уровне анализа флор районов, у Камчатки образуются с другими территориями Циркумбореальной области. Количество восточноазиатских видов настолько мало, что это почти не находит отражения в спектрах элементов флоры и типов ареала. Небольшое влияние оказывает увеличение количества приокеанических таксонов на юге Камчатки и островах, что выражается в существовании отчетливых географических связей с другими островными территориями северной Пацифики.
Здесь же уместно обсудить вопрос о восточно-североамериканских связях флоры Камчатки. Выяснено (Qian et al., 2003а), что количество видов сосудистых растений, имеющих такой ареал, возрастает при продвижении с севера на юг и достигает максимума на 30±5° с.ш. Указанная закономерность, возможно, свойственна и печеночникам. По крайней мере, количество таких таксонов относительно велико в Японии и Корее, но в Гонконге, на 23° с.ш., предстаивтели с этим типом ареала уже вообще не известны (Li, Corlett, 2003). В нашем регионе восточноазиатско−восточно-североамериканских видов нет. В целом оказывается низким и количество сугубо амфипацифических видов, их всего 6: Gymnomitrion pacificum, Macrodiplophyllum plicatum, Ptilidium californicum, Cephalozia otaruensis, Macrodiplophyllum imbricatum, Schofieldia monticola.
Специфичность флор выделенных районов Камчатки на родовом уровне проявлена следующим образом. Родов, известных только из одного района - 9. Это разнородная группа, каждый член которой показывает определенные связи как Камчатки в целом, так и специфику самого флористического выдела. Южные связи прослеживаются на уровне 4 родов из Восточной Камчатки (Targionia, Ricciocarpos, Geocalyx, Crossogyna), 1 - с Северной (Anthoceros) и 1 - с Командорских островов (Porella). Амфипацифические связи представлены единственным родом Schofieldia, обнаруженным на севере полуострова. Изолированные местонахождения еще двух родов (Mannia, Herbertus) объясняются редкостью подходящих субстратов на полуострове или общим реликтовым характером распространения их представителей.
Отчасти специфику флоры отражают роды, известные в сопредельных регионах, но не найденные на Камчатке. Таких родов оказалось 20. Лишь один их них - монотипный Mesoptychia - встречается на Чукотке, но не известен на Камчатке. 13 родов (Alobiellopsis, Cololejeunea, Crossocalyx, Iwatsukia, Lejeunea, Liochlaena, Neohattoria, Nipponolejeunea, Nowellia, Pallavicinia, Pedinophyllum, Reboulia, Trichocolea) распространены на Южных Курилах. Четыре рода (Acrobolbus, Anastrepta, Douinia, Gyrothyra) отмечены на Алеутах и еще 2 (Metzgeria, Metacalypogeia) - как на Алеутах, так и Южных Курилах.
Следует отметить три обстоятельства: 1) все южно-курильские роды, за исключением Alobiellopsis, Neohattoria и Nipponolejeunea, встречаются и в Северной Америке; 2) все алеутские роды, за исключением Gyrothyra, известны в Восточной Азии; 3) несомненно реликтовый характер носит распространение на Алеутах Metacalypogeia и Acrobolbus, не встреченных на Камчатке. Иными словами, отсутствие ряда родов на Камчатке объясняется не климатическими и миграционными причинами, а третьими, среди которых важнейшим фактором является подавляющее действие активного вулканизма, сопровождающего почти все районы изученного региона на всем протяжении неогена.
Использование корреляционного анализа родовых спектров по флористическим районам Камчатки и сопредельных выделов показало при нижнем пороге коэффициента корреляции в 90% образование 4 простых связей, причем можно предположить причины их взаимоотношений. Центральная Камчатка связывается с Командорами - оба района обладают хорошо выраженным поясом арктических пустошей и горных тундр, не нарушенных современным вулканизмом. Восточная Камчатка оказывается родственной Северным Курилам - оба района характеризуются высокой современной вулканической активностью. Связь Восточной Камчатки с Восточной Чукоткой объясняется океаничностью обоих районов при относительно суровом климате. Командоры связаны с Северными Курилами как два района с более теплым, чем на Чукотском п-ове и Восточной Камчатке, океаническим климатом.
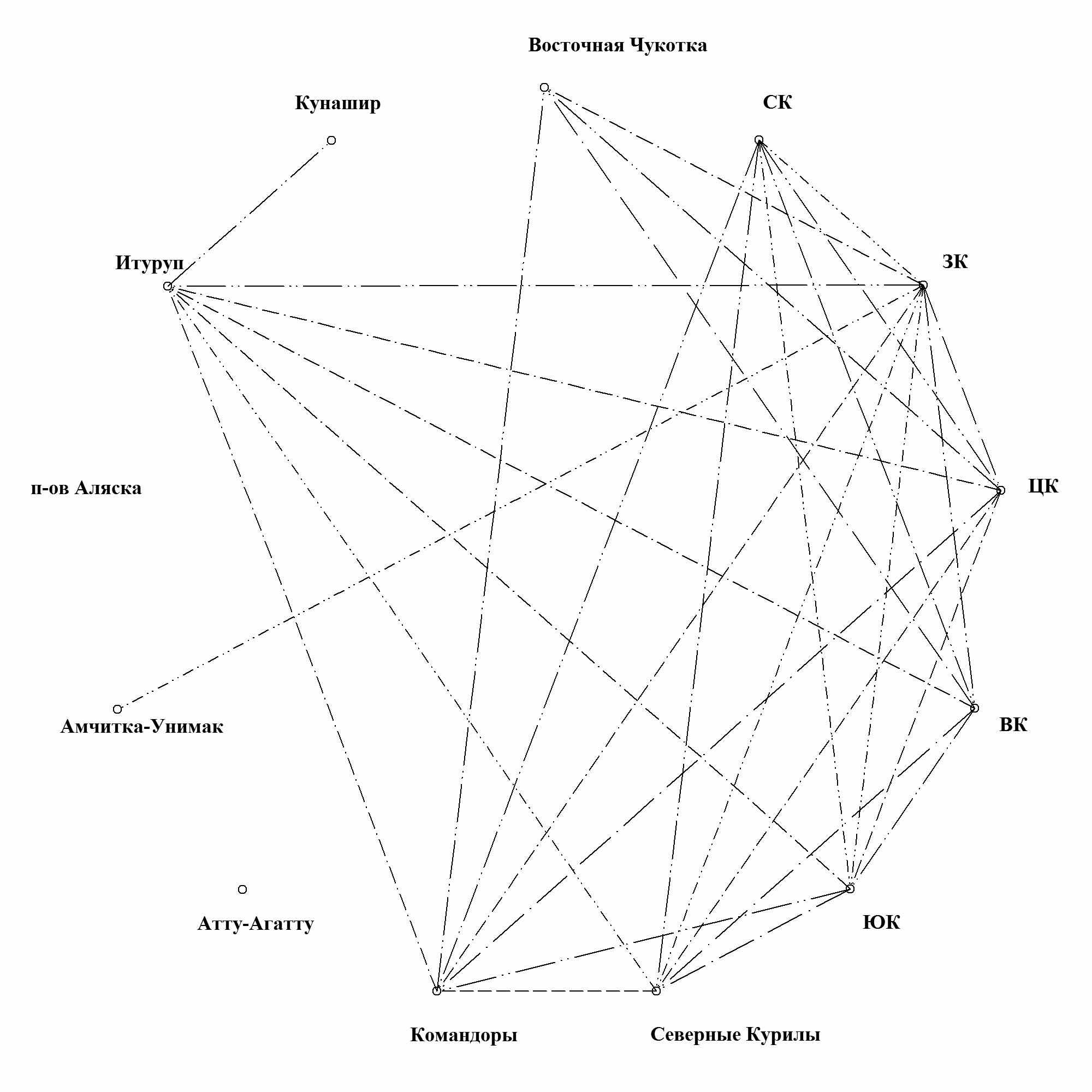 |
| Рис. 9. Корреляция между таксономическими родовыми спектрами районов, при нижней границе коэффициента 80%. |
На видовом уровне, кроме экспертной оценки специфичности каждого флористического подразделения Камчатки, было проведено сравнение районных флор, расположенных по широтному и долготному градиентам по количеству общих видов (Рис. 10, 11).
При сравнении Центральной Камчатки с районами, расположенными по широтному градиенту, наиболее родственной ей территорией оказывается не другие районы Камчатки, а Восточная Чукотка (93 общих таксона). Возможно, это является следствием чрезвычайно суровых природных условий в верхних поясах гор Центральной Камчатки, сходных с таковыми на Чукотке, а также относительно сухим климатом, выражающимся в разнообразии мезо- и криоксерофитных таксонов. Родственными Центральной Камчатке оказываются другие районы полуострова, а также значительно южнее расположенный о-в Кунашир (75 общих таксонов). Связи с последним объясняются, во-первых, наличием ряда теплолюбивых реликтов в нижних поясах гор Срединного хребта (Bazzania), широко распространенных на Кунашире, а также в группе арктических монтанных видов, обычных на Камчатке и встречающихся на Кунашире на лавовых потоках на вершине вулкана Тятя (выше 1500 м над ур. м.). При изучении родства выделов, расположенных по условной линии "запад-восток", к Центральной Камчатке оказываются наиболее близки Восточная Камчатка (98 таксонов) и Командоры (90), расположенные на едином миграционном пути Hultenia и ближайшие территориально. Вне нашего региона тесные связи (но значительно меньшие) с Центральной Камчаткой показывают флоры северо-западной Карелии (Regio Kuusamoensis, 80 общих таксонов) и восточная Якутия (76). В западном полушарии наибольшее количество общих видов зарегистрировано с Ньюфаундлендом (67). Сходные тенденции прослеживаются и при сравнении флоры Восточной Камчатки.
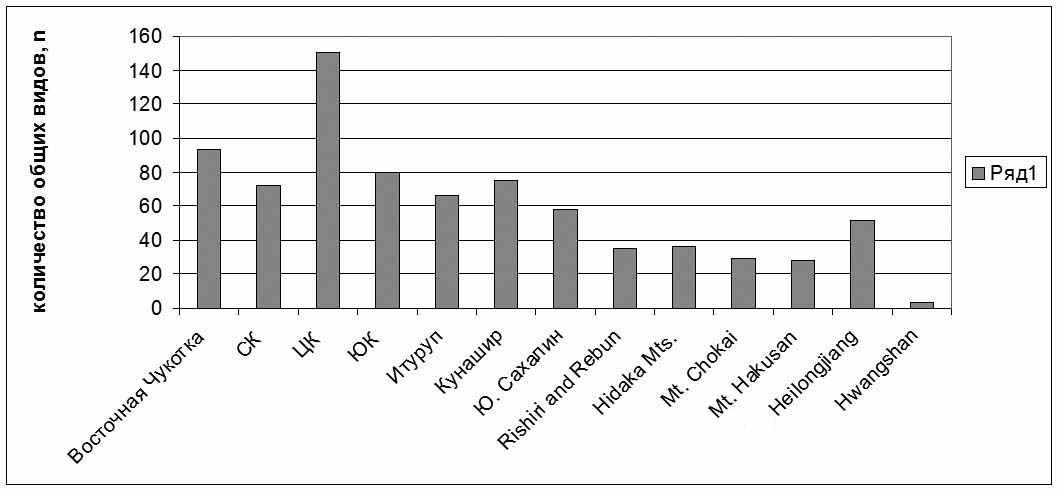 |
| Рис. 10. Изменение количества общих видов в районах по сравнению с флорой Центральной Камчатки по широтному градиенту. |
При изучении флористических списков, составленных по каждому району Камчатки и прилегающих островов, выяснилось, что во всех районах встречается 27 видов, в 6 районах - 26, в 5 - 25, в 4 - 19, в 3 - 36, в 2 - 33 и, наконец, только в 1 районе - 67 видов.
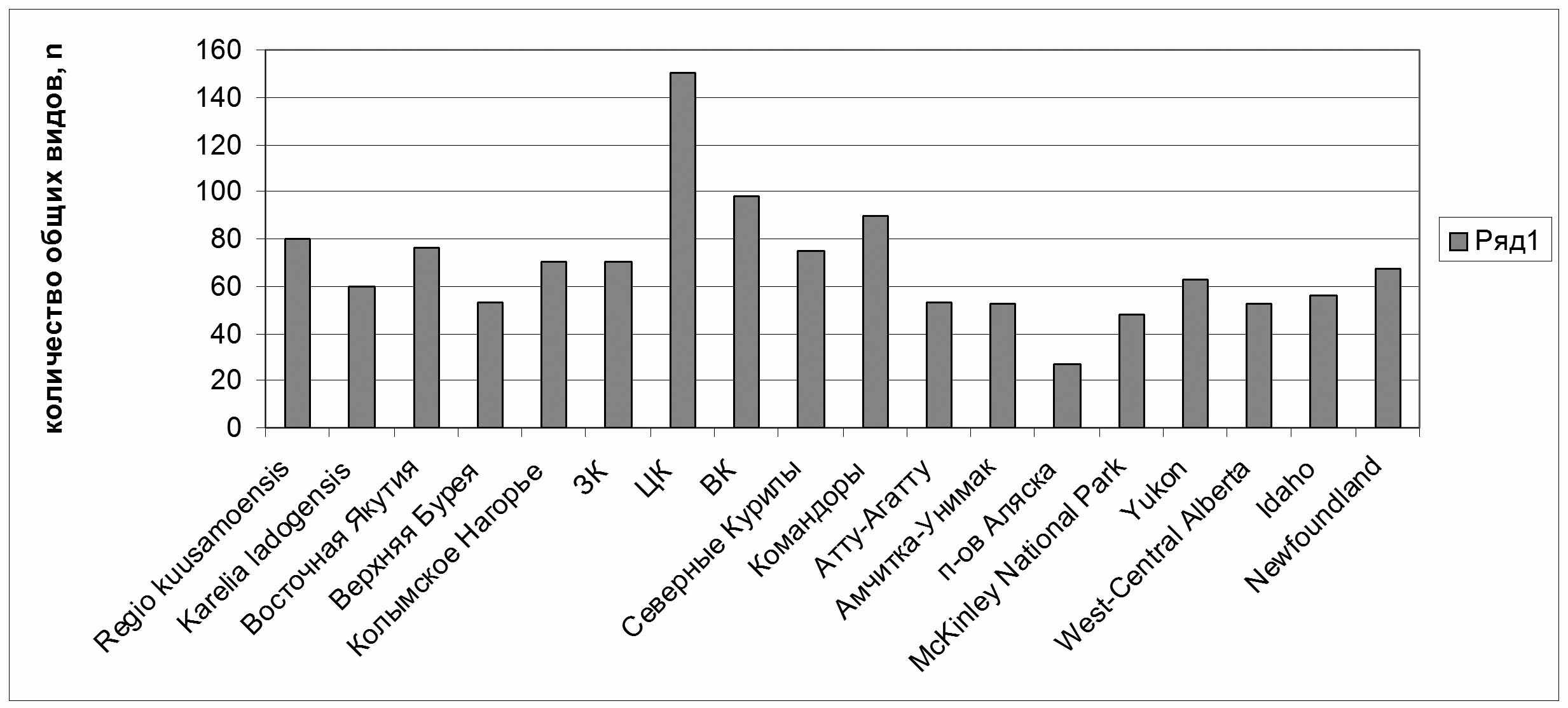 |
| Рис. 11. Изменение количества общих видов в районах по сравнению с флорой Центральной Камчатки по долготному градиенту. |
Если проследить какими видами в изученном регионе связываются только 2 района, выясняется, что наиболее четкие связи образуются между Центральной Камчаткой и соседними районами: Северной, Восточной и Южной Камчаткой. Как и можно было предположить, связь Северной и Центральной Камчатки прослеживается на уровне таксонов северного распространения: Solenostoma sphaerocarpum, Solenostoma jenseniana, Lophozia propagulifera, Blepharostoma trichophyllum var. brevirete, Cryptocolea imbricata. Восточная и Центральная Камчатка связаны между собой, во-первых, таксонами, распространение которых на Камчатке приурочено только к термальным источникам (Solenostoma fusiforme, Fossombronia) и, во-вторых, с видами, встречающимися в горных тундрах северного полушария (Sphenolobus saxicola, Schistochilopsis grandiretis, Scapania brevicaulis). Южная и Центральная Камчатка связаны между собой видами, камчатские нахождения которых лежат вблизи северной границы распространения в Восточной Азии: Obtusifolium obtusum, Marsupella funckii, Lepidozia reptans, Nardia unispiralis, причем последний вообще встречается только в островной Пацифике (от Японии до Камчатки). Выделяется также группа с восточнокамчатско-южнокамчатским распространением. Таких видов насчитывается 4. Это - восточноазиатские Plectocolea vulcanicola и Nardia assamica, из них общее распространение первого связано исключительно с термальными источниками, а также циркумполярные, вероятно, случайные виды Pellia epiphylla и Diplophyllum obtusifolium.
Пятью видами маркируется камчатский участок трансокеанского пути Азия - Америка (Северные Курилы, Южная Камчатка и Командоры). Только в них известно 5 видов: Anthelia julacea, Moerckia blyttii, Nardia compressa, Orthocaulis floerkei и Plectocolea infusca var. ovalifolia. Как видно, этот путь не затрагивает Восточную Камчатку, ближайшую к Командорам. Вероятно, последнее является следствием новейшего активного вулканизма на большой части востока полуострова, уничтожающего промежуточные звенья азиатско-американских ареалов.
Следует отметить два противоположных, но закономерных, результата проведенного в этом разделе анализа, которые можно выразить в общем виде положением Аристотеля о единстве противоположностей. Каждый район своеобразен и обладает присущими только ему специфическими чертами (набор видов, ареалогическая характеристика, генетические связи и т.п.), но вместе с тем, этот конгломерат четко отграничен от прилежащих районов, что "при удалении" формируется впечатление единства всех районов и их отличия от прилежащих. Таким образом, флора печеночников Камчатки и прилегающих островов, несмотря на наличие локальных девиаций, заслуживает выделения в самостоятельную единую флористическую провинцию.
Глава 8. Фитогеографические связи Камчатки и прилегающих островов через Курильскую и Алеутскую островные дуги
При рассмотрении флорогенеза нашего региона, очевидно, что сначала флора пра-Камчатки была термофильная и связывалась сухопутным путем с юго-восточной Азией. На границе миоцена и плиоцена произошло важнейшее геоморфологическое событие: объединение пра-Камчатки с северо-восточной Азией (Шанцер, 1974). Последнее открыло на полуостров путь массам северо-восточноазиатских видов (при этом следует помнить, что сама флора континентальной северо-восточной Азии была принципиально отличной от современной). Так, при крепнущих северных связях ослабевали южные и очень ограниченную роль играли восточные. Прогрессирующее похолодание, чреда оледенений в плейстоцене, вероятно, почти начисто уничтожили древнюю теплолюбивую флору. Образовавшаяся "пустота" была заполнена видами, распространенными на территории современного Корякского нагорья. Затрудненность миграций по островным дугам привела к "осевернению" современной флоры Камчатки, по сравнению с регионами азиатской России, расположенными на сходных широтах. Наиболее ярко это проявилось в неспецифичности комплексов темнохвойных лесов. Вместе с тем, Камчатка обладает определенным своеобразием, показывающим ее восточные и южные связи. Эти отличительные черты могут трактоваться как реликтовый остаток древней флоры, но могут рассматриваться и как результат плейстоцен-голоценовой миграции. В пользу последнего говорят факты современного распространения ряда трансалеутских видов, которые не могли пройти по северному сухопутному мосту - Берингии - во время регрессий вследствие их выраженной термофильности. Так или иначе, целесообразно обсудить современные фитогеографические связи Камчатки, осуществляющиеся через Курильские и Алеутские острова.
При всем разнообразии северопацифических частей ареалов видов печеночников, охватывающих как Азию, так и Америку, даже при беглом анализе, выявляется три группы таксонов, различающиеся распространением. У одних азиатские и американские части ареала связываются через Берингию, у других – через Алеутские острова. Виды, свойственные каждой из групп, весьма значительно различаются по экологическим предпочтениям. Это или арктические виды, как Mesoptychia sahlbergii Lindb., распространенный в арктической Пацифике, или бореальные и неморальные как представители родов Porella и Radula. Кроме того, остается еще одна, наиболее крупная группа таксонов, распространенных как в районе Берингии, так и на Алеутской дуге. Происходившие гляциоэвстатические трансгрессии и регрессии моря объединяли и разъединяли отдельные острова Алеут, но никогда не приводили к образованию сплошного моста суши "Камчатка - Аляска".
 |
| Рис. 12. Распространение Metacalypogeia cordifolia (Steph.) H. Inoue в Северной Пацифике. |
При детальном рассмотрении списка Алеутских печеночников, не встречающихся на Камчатке (или северной Аляске), но распространенных на противоположной оконечности Алеутской дуги, выясняется, что Камчатку с Алеутами роднят, в основном, восточноазиатские таксоны (Lophozia lacerata, L. silvicoloides, Conocephalum japonicum, Jungermannia exertifolia и др.) и северные (Jungermannia polaris, Marsupella boeckii, Nardia breidleri, Peltolepis quadrata, Scapania obscura и др.). В свою очередь, Северную Америку с Алеутами связывают западно-американские или преимущественно атлантические таксоны (Douinia ovata, Gyrothyra underwoodiana, Macrodiplophyllum imbricatum и др.), а также теплолюбивые представители родов Plagiochila, Radula и Porella. Причем интересно, что практически все представители родов Radula и Porella распространенные на Алеутах, но не встречающиеся на Камчатке, в дальнейшем появляются на Южных Курилах и Японском архипелаге. Вероятно, в данном случае имеет место вторичный разрыв ареала, обусловленный изменениями климата в конце Плейстоцена, либо высокой вулканической активностью на Курилах.
Важное фитогеографическое значение имеет линия Татеваки (Tatewaki-line), отделяющая Командоры от Алеут в узком смысле (Tatewaki, 1963b). Известными на Командорах, но не найденными на остальной части Алеутской гряды, оказываются 43 вида, из них один (Riccardia aeruginosa) не встречается на Камчатке, а 17 – на Аляске. Известны на Алеутах, в узком смысле, но не встречены на Командорах – 38 видов, из которых на Аляске не найдены Metacalypogeia cordifolia и Acrobolbus ciliatus, а на Камчатке - 21 таксон. Для 4 восточноазиатских видов (Conocephalum japonicum, Lophozia lacerata, Plectocolea infusca, Riccardia aeruginosa) Командоры – форпост распространения в восточном направлении. Для 5 североамериканских видов (Gyrothyra underwoodiana, Plagiochila schofieldiana, P. corniculata, P. semidecurrens, Radula polyclada) Алеуты, в узком смысле, являются границей в распространении на запад.
Курильская гряда, вытянувшаяся на 1200 км в северо-восточном направлении от берегов о-ва Хоккайдо к южной оконечности п-ова Камчатка, представляет собой гигантский океанический мост между самобытными флорами этих территорий. Не подлежит сомнению существование огромной разницы между субарктической флорой нижних высотных поясов южной Камчатки и неморальной, с большим количеством субтропических видов, флорой долин и побережий о-ва Хоккайдо. По Курильской островной дуге проходит граница между Циркумбореальной и Восточноазиатской флористическими областями. Многочисленные классификации флористического покрова на Курилах описаны В.Ю. Баркаловым (2002), однако все они были построены на основе анализа распространения сосудистых растений.
С целью сравнения были проанализированы флоры печеночников о-вов Кунашир, Итуруп, а также Северных Курил, по которым более или менее достоверные данные. Для сравнения в анализ были вовлечены сопредельные регионы различного размера, но имеющие сходную орографию и степень изученности, и, как следствие, близкие показатели известного таксономического разнообразия: Чукотский автономный округ (далее Чукотка), п-ов Камчатка, о-в Хоккайдо и горный массив Chichibu-Okutama в средней части о-ва Хонсю.
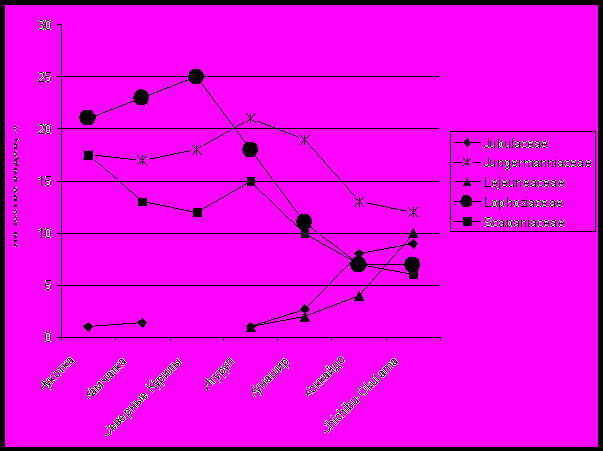 |
| Рис. 13. Изменение вклада в формирование региональных флор отдельных семейств. |
Корреляционный анализ родовых спектров приводит к образованию двух клад, одна из которых связывает все регионы от Чукотки до Кунашира, вторая объединяет о-в Хоккайдо с массивом Chichibu-Okutama. Между двумя массивами достоверная корреляция отсутствует. При этом Кунашир достоверно связан только с ближайшим о-вом Итуруп и с п-овом Камчатка, а Чукотка образует достоверные связи с Камчаткой, Северными Курилами и Итурупом.
Степень сходства сравниваемых северо-западно-пацифических флор может быть измерена числом общих таксонов, а характер, или природу связей позволяет выявить географическая характеристика связующих видов. Общими для всех территорий, вовлеченных в анализ (Чукотка, п-ов Камчатка, Северные Курилы, о-ва Итуруп, Кунашир, Хоккайдо, массив Chichibu-Okutama) являются 17 видов. Среди них преобладают арктические и арктобореальные (14 таксонов), а также присутствуют мультизональные (3 вида), причем, среди последних один - восточноазиатский Conocephalum japonicum - визитная карточка флоры печеночников Восточной Азии, от севера до юга. Таким образом, взаимосвязи флор осуществляются, главным образом, через группу видов, произрастающих на севере в равнинных и горных тундрах, а на юге - в высокогорьях.
При сокращении числа флор (вышеуказанные, но без Чукотки) вовлеченных в анализ, в группу общих попадают также три бореальных вида (Conocephalum conicum, Pellia endiviifolia, Ptilidium pulcherrimum. Общими для всех подразделений Курил являются 46 видов.
 |
| Рис. 14. Изменение участия видов, относящихся к избранным широтным элементам в сравниваемых флорах. |
Неморальное веяние в составе общих видов флоры начинает проявляться с Итурупа и становится очевидным с Кунашира и южнее. Так, Кунашир роднят с Хоккайдо 93 таксона (более половины из обеих флор). Причем наряду с большим числом арктических, арктобореальных и бореальных таксонов (всего 57), б