
Флора и фитогеография печеночников (marchantiophyta, anthocerotophyta) камчатки и прилегающих островов 03. 00. 05 ботаника
| Вид материала | Автореферат диссертации |
- Флора и растительность копаней ярославской области 03. 00. 05 ботаника, 463.62kb.
- Сохранение биоразнообразия камчатки и прилегающих морей южно-Камчатский заказник: изменения, 45.7kb.
- Гидрофильная флора сосудистых растений как модельный объект для инвентаризации и анализа, 555.67kb.
- На семинаре предполагается рассмотреть следующие основные темы, 47.38kb.
- Кузьмицкого Алексея Алексеевича, формирование по сути однопартийного представительного, 1278.31kb.
- И с Россией Андреевский флаг, 457.61kb.
- Соломон Шульман, 2770.57kb.
- Географическое положение, 122.67kb.
- Дальнего Востока России. Официальный язык конференции русский доклад, 45.87kb.
- Флора Вафина (творческий псевдоним-Флора) автор слов и музыки, исполнитель романсов,, 133.03kb.
Начало изучения печеночников Камчатки было положено доктором Горфратом Тилезиусом (H. Tilesius) – участником кругосветной экспедиции под руководством И.Ф. Крузенштерна. Им было собрано 8 видов, определенных и опубликованных G. Wahlenberg (1811). К сожалению, ни в статье, ни в этикетках, прилагаемых к материалу, не отражены условия произрастания и места сбора. Остается только предполагать, что коллекция была собрана в окрестностях Авачинской бухты, посещенной экспедицией Крузенштерна. Это обстоятельство тем более печально, что с тех пор не были найдены два вида из списка Wahlenberg Asterella saccata и Targionia hypophylla.
После упомянутой работы публикации были немногочисленны и базировались, в основном, на сборах неспециалистов (в лучшем случае, ботаников, занимающихся сосудистыми растениями). К 1970 году, за 170-летнюю историю исследования, для Камчатки указывалось около 50 таксонов печеночников. Последнее привело к исключительной по своей неверности фразе в работе H. Persson, о том, что «число известных с Камчатки мохообразных впоследствии увеличится на 30-40%» (Persson, 1970: 209). Исследования последних 10 лет увеличили это число не на 30-40%, а на 340%. Чрезвычайная активность проведения исследований в последние годы объясняется целым рядом причин.
В 2003 году мной была предпринята попытка синтеза уже накопленных к тому времени данных и опубликован "Предварительный список печеночников полуострова Камчатка" (Bakalin, 2003d). К тому времени для Камчатки указывалось 80 видов печеночников. После обработки собственных сборов 2001 и 2002 гг. мне удалось пополнить этот список еще 78 видами. Высокая интенсивность проведения исследований в последующие годы (Потемкин, 2003 и др.) вызвала необходимость издания значительного списка дополнений к флоре Камчатки, а также обзора новейших публикаций, содержащих сведения по флоре печеночников Камчатки. Были опубликованы данные о распространении 150 видов печеночников по территории полуострова, в том числе 33 указано для Камчатки впервые (Bakalin, 2005a). С учетом этих новых данных, а также на основании определения недавних коллекций количество видов на п-ове Камчатка и прилегающих островах достигло 227. Число это, по-видимому, не является окончательным и еще не менее 10 таксонов (5%) может быть обнаружено в регионе.
Глава 4. Растительные сообщества региона и участие печеночников в их сложении
История развития флоры Камчатки подразделяется на два этапа: от позднего мела до начала плиоцена и постплиоценовый. Самые ранние из известных отложений (Челебаева, 1971) относятся к позднему палеоцену или раннему эоцену. В это время во флоре присутствуют как представители неморальных (Acer, Tilia, Ulmus), так и субтропических родов (Dennsteadtia, Glyptostrobus, Metasequoia, Taxodium, Magnolia). Дальнейшее развитие флоры на пра-Камчатке свидетельствует о постепенном, но неуклонном и прогрессирующем изменении в сторону отбора холодостойких пород и сопряженного с этим видообразования (ср. Tiffney, 1985). В течение плейстоцена происходило колебание, преимущественно между площадью, занимаемой сообществами различных типов, но не выражающееся в кардинальных изменениях флористического состава. Можно предположить, что флора печеночников с тех времен сохранилась почти в неизменном виде, а происходившая трансформация имела негативный характер и проявлялась в уничтожении популяций и отдельных видов целиком вследствие активного вулканизма.
Очерк растительности региона с описанием участия печеночников в сложении флоры. Наиболее разнообразны природные сообщества на п-ове Камчатка. Полный высотный спектр встречается в северной части Центральной Камчатской депрессии. Здесь последовательно сменяются ельники, лиственничники, каменноберезняки, ольховники и кедровники и, наконец, горные тундры и альпийские пустоши. Вблизи побережий, на севере и юге п-ова хвойные леса не встречаются. На западе п-ова широкая полоса вблизи побережья занята приморскими кочкарными тундрами и маломощными болотами, в удалении появляются ольховники, кедровники и березняки. В других районах растительность вблизи побережий представлена сочетанием ольховников, березняков и, значительно реже, кедровников. В долинах более или менее крупных рек, при некотором отдалении от побережья, развиты тополевники, ивняки, чозениевые, черемуховые и ольховые леса. Болота не имеют значительного распространения в регионе. Наибольшие их массивы сосредоточены на западном побережье и в северной части п-ова, где они часто неотличимы от влажных тундр. По всей территории широко распространены явления высотно-поясной инверсии.
При рассмотрении высотной поясности различных горных районов азиатской России, выявляется два комплекса специфичных видов: первые из них приурочены к нижнему высотному, таежному, поясу, вторые к верхнему, тундровому. Переходные сообщества (криволесья, к которым по физиономическим характеристикам должны быть отнесены каменноберезняки и стланиковые сообщества) обычно бедны во флористическом отношении. Камчатка представляет собой исключение из этой закономерности. Площадь хвойных лесов сильно сокращалась во времена плейстоценовых оледенений, и после отступления ледников возвращение этих лесов в прежние границы происходило из небольших по площади рефугиумов, а не широким фронтом, как в большинстве районов северной Евразии, что привело к уменьшению количества связанных с хвойными лесами видов. Только в каменноберезняках встречается 8 видов (Herbertus aduncus, Bazzania bidentula, B. ovifolia, Anastrophyllum michauxii, Tritomaria exsecta, Scapania scandica var. scandica, Frullania davurica, Calycularia crispula). Семь из них находятся на северном пределе своего ареала в Восточной Азии. Один (Herbertus aduncus) характеризуется реликтовым распространением. Кроме того, к каменноберезнякам, несомненно, тяготеют еще два вида. Так, Lophozia ascendens, распространен повсюду в Голарктике только в хвойных лесах, и лишь на Камчатке этот вид встречается в каменноберезняках и пойменных тополевниках. Второй вид, Radula constricta, произрастает Восточноазиатской флористической области, а вне ее обнаружен только на Камчатке, где встречается в каменноберезовых и ольховостланиковых сообществах.
Генетическая связь перечисленных специфичных видов с сообществами березняков отсутствует. Возможно, что во время температурных колебаний климата эти виды сохранились в относительно широко распространенных, даже в периоды оледенений, каменноберезняках, и в настоящее время остались верны этому типу леса. Так или иначе, эта фракция специфичных таксонов каменноберезняков составляет большую часть видов с южными связями Камчатской флоры печеночников.
Тундровые сообщества в регионе занимают около 1/5 территории и именно к ним приурочено основное разнообразие печеночников. Выделяются две группы тундр по высотному положению: 1) долинные и приморские, чье существование обусловлено термическими инверсиями, ветровым режимом и охлаждающим действием океана; 2) горные, обусловленные низкими температурами и, отчасти, ветровым режимом. При значительном, на первый взгляд, почти полном физиономическом сходстве основное разнообразие печеночников сосредоточено все-таки в горных тундрах. Только в тундрах обнаружено 80 видов.
Современный вулканизм и ледниковая деятельность приводит к существенному обеднению флористических комплексов, подвергающихся прямому их воздействию, и в таких районах отсутствует ряд широко распространенных в горных тундрах Камчатки видов, которые не способны к активному и постоянному генеративному и вегетативному размножению. Однако своеобразие флоры в таких районах несколько повышается за счет присутствия таксонов, занимающих молодые субстраты.
Во флористических группировках окрестностей термальных источников обнаружен 41 вид печеночников. Лидером по терморезистентности оказался Marchantia polymorpha L. Интерес представляют виды, обнаруженные на Камчатке только в окрестностях термальных водотоков. Как выяснилось они не образуют единого комплекса, а являются разнородной группой (Calypogeia neogaea, Anthoceros agrestis, Riccia huebeneriana, Fossombronia sp., Plectocolea vulcanicola, Solenostoma fusiforme, Leiocolea bantriensis).
По типам местообитаний наиболее богатыми являются переувлажненные участки в горных тундрах, нивальные сообщества, каменистые берега в истоках ручьев. В нижерасположенных поясах основное разнообразие привязано к кристаллическим субстратам - скалам, останцам, речным каньонам и др.
Глава 5. Конспект флоры печеночников Камчатки и прилегающих островов
Расположение семейств в конспекте приведено согласно системе R. Grolle в Nomina generica Hepaticarum (1983), за исключением подсемейства Lophozioideae, которое рассматривается как самостоятельное семейство Lophoziaceae, и перемещения Scapaniaceae непосредственно следом за Lophoziaceae. Род Leiocolea помещается в Jungermanniaceae. Виды и роды внутри семейства размещены в алфавитном порядке, а их таксономическая трактовка соответствует принятой мной ранее (Bakalin, 2003d, 2005а). Сокращения авторов таксонов даны согласно списку R.K. Brummit и C.E. Powel (1992).
По каждому виду приведены отдельные аннотации (для п-ова Камчатка, Северных Курил и Командор), составленные по следующей схеме: 1) присутствие в образцах, изученных автором, органов вегетативного и генеративного размножения по следующей шкале: часто – структура выявлена с более чем в 50% образцов, изредка – 20-50%, редко – менее 20%; если имеется указание на постоянное присутствие органов размножения, то оно говорит о том, что такие структуры выявлены хотя бы у одного растения, но из каждого изученного образца; 2) распространение по территории полуострова и групп островов; 3) экологические условия произрастания; 4) сопутствующие таксоны; 5) высота над уровнем моря местонахождений вида.
 |
| Рис. 2. Флористическое районирование Камчатки и прилегающих островов. |
Для некоторых видов добавлены комментарии таксономического, экологического, географического или иного содержания. Для каждого таксона приведена карта, показывающая его местонахождения на Камчатке и прилегающих островах. В тех случаях, если образец вида не был изучен автором, а в литературных источниках не указана экология, условия произрастания даны предположительно в квадратных скобках.
Ниже приведен список родов печеночников, обнаруженных во флоре Камчатки и прилегающих островов. В скобках после родового названия указано число видов в роде.
ANTHOCEROTOPHYTA
Anthocerotaceae: Anthoceros (1).
MARCHANTIOPHYTA
Herbertaceae: Herbertus (1).
Pseudolepicoleaceae: Blepharostoma (1).
Lepidoziaceae: Bazzania (3), Kurzia (1), Lepidozia (1).
Calypogeiaceae: Calypogeia (6).
Cephaloziaceae: Cephalozia (9), Cladopodiella (2), Hygrobiella (1), Odontoschisma (2), Pleurocladula (1), Schofieldia (1).
Cephaloziellaceae: Cephaloziella (10).
Antheliaceae: Anthelia (2).
Jungermanniaceae: Crossogyna (1), Cryptocolea (1), Cryptocoleopsis (1), Jungermannia (6), Leiocolea (4), Mylia (1), Nardia (9), Plectocolea (6), Solenostoma (7).
Lophoziaceae: Anastrophyllum (2), Barbilophozia (4), Gymnocolea (1), Isopaches (2), Lophozia (16), Obtusifolium (1), Orthocaulis (6), Protolophozia (1), Saccobasis (1), Schistochilopsis (5), Sphenolobus (3), Tetralophozia (1), Tritomaria (3).
Scapaniaceae: Diplophyllum (4), Macrodiplophyllum (3), Scapania (23).
Gymnomitriaceae: Eremonotus (1), Gymnomitrion (4), Marsupella (12), Prasanthus (1).
Geocalycaceae: Chiloscyphus (4), Harpanthus (1), Lophocolea (3)
Plagiochilaceae: Plagiochila (2).
Radulaceae: Radula (3).
Ptilidiaceae: Ptilidium (3).
Porellaceae: Porella (1).
Frullaniaceae: Frullania (3).
Codoniaceae: Fossombronia (?1).
Pelliaceae: Pellia (3).
Allisoniaceae: Calycularia (2).
Pallaviciniaceae: Moerckia (1).
Blasiaceae: Blasia (1).
Aneuraceae: Aneura (1), Riccardia (4).
Metzgeriaceae: Apometzgeria (1), Metzgeria (1).
Targioniaceae: Targionia (1).
Conocephalaceae: Conocephalum (2).
Aytoniaceae: Asterella (2), Mannia (1).
Cleveaceae: Athalamia (1), Peltolepis (1), Sauteria (1).
Marchantiaceae: Marchantia (3), Preissia (1).
Ricciaceae: Riccia (4), Ricciocarpos (1).
Глава 6. Общие черты и генетические связи флоры печеночников Камчатки и прилегающих островов
В современной и более старой литературе таксономический и географический анализы проводятся раздельно (часто занимая в монографиях даже разные главы). На самом деле, если речь идет действительно об анализе флоры, а не о констатации отдельных фактов (как количество видов принадлежащих к определенному элементу флоры или семейству), результаты осмысления таксономических спектров начинают приобретать географический оттенок. Таким образом, по сути, оба типа анализа сливаются и служат достижению единой цели. Осознание этого факта приводит к необходимости совместного рассмотрения, или даже некого объединения географической и таксономической характеристик каждой флоры.
Таксономический и ареалогический анализы, являются ключом для познания флористического феномена конкретной территории. Оба типа анализа преследуют, в сущности, одни цели: отобразить специфику флоры, ее генетические связи, дать достоверный источник для реконструкции флорогенеза. Однако они проводятся на основании как бы разных срезов одного массива. Оба метода дают в качестве конечного результата факты, демонстрирующие сходство или несходство ареалов таксонов или таксономического состава сравниваемых флор.
В проведенный анализ (как таксономический, так и географический) включены не только виды, но и некоторые внутривидовые таксоны, в частности разновидности отдельных видов, рассматривающиеся в анализе как самостоятельные единицы. Сделано это по двум причинам. Во-первых, ранг таксона у печеночников (разновидность, подвид или вид) в некоторых случаях (если не в большинстве) определить очень трудно вследствие недостаточной изученности морфологической вариабельности, генетических барьеров, экологии и распространения. Во-вторых, географические связи и характерные черты флоры отражаются иногда не общим распространением вида, а присутствующими на территории разновидностями.
Для получения впечатления о характере флоры печеночников достаточно использования показателей, рассчитанных только по трем ведущим семействам. Как показывают аналитические исследования региональных флор северной Голарктики (Арктики и Гипоарктики, в смысле Б.А. Юрцева, 1966), проведенные Н.А. Константиновой (1998), три ведущих семейства включают более половины видов всей флоры. Указанная закономерность делает ненужным (а часто даже и вредным, из-за создания помех случайного характера) проводить при характеристике флоры анализ 10 ведущих семейств, как это делается (и вполне оправдано) у сосудистых растений.
В основе географического анализа лежит схема классификации типов распространения принятая Н.А. Константиновой (2000), но с некоторыми изменениями. Наиболее существенное их них касается группы видов, связанных с горами. Традиционно при описании приуроченности в распространении к горам в название ареала вводилось слово монтанный, или альпийский (аркто-альпийский, борео-монтанный и т.п.). Некоторые авторы также выделяли так называемые горные виды в качестве самостоятельного элемента флоры. Под ними обычно подразумевались горные виды, не заходящие в Арктику. Последнее положение неоднократно подвергалось критике в советской литературе. Однако, если даже полностью отказаться от выделения монтанного географического элемента, проблема, по всей вероятности, будет решена лишь частично, поскольку в названиях других элементов будут присутствовать сочетания чуждых друг другу по смыслу слов (аркто-альпийский, гипоарктически-горный и т.п.). Очевидно, что термин горный или альпийский обозначает приуроченность видов к орографическим системам и не имеет широтной окраски. Выход из этой ситуации возможен при «вынесении за скобки» показателя монтанности.
При проведении сравнительного анализа сопоставлялись таксономические и географические спектры флоры печеночников Камчатки в целом, с другими регионами, так и флористических районов в ее составе с другими небольшими флорами. Для сравнения была проведена процедура отбора флор разного ранга (по литературным данным и собственным материалам) и условное разделение их на категории районных и региональных. К сожалению, при этом не удалось применить простой и логичный принцип: флоры регионов занимают большую площадь и богаче таксономически, чем флоры районов. Во многих случаях из-за недостаточной изученности разнообразия печеночников для огромных по площади территорий приводятся весьма скудные списки видов. Это особенно касается ряда северных притихоокеанских флор, сравнение с которыми было необходимым. В таких случаях основным принципом разделения являлся уровень флористического богатства. Результатом применения такого подхода явились существенные перекосы. Так, например, Колымское нагорье, имеющее площадь в 3 раза превышающую площадь Камчатки, было отнесено к районам, поскольку имеет, по опубликованным данным, таксономическое разнообразие в 91 таксон. Очевидны два обстоятельства: 1) приводимая цифра отражает таксономическое разнообразие не более чем на 60%, 2) проводить прямое сравнение флоры нагорья с флорой Камчатки и прилегающих островов неправильно, поскольку в таком случае нагорье полностью включится в Камчатку.
Условная граница для разделения флор на районные и региональные была определена на уровне приблизительно в 150 видов. Применение одной меры для всех я считаю вполне оправданным. Ignatov (1993) показал, что на территории бывшего СССР таксономическое разнообразие мхов существенно повышается в горных и приокеанических районах, в то время как разнообразие сосудистых растений возрастает практически везде пропорционально уменьшению широты. Закономерности изменения регионального таксономического разнообразия мхов на территории бывшего СССР действительны и для печеночников, а с некоторыми коррективами, по-видимому, вообще по всему земному шару. Крупные флоры, расположенные на разной широте, от субарктики до южно-умеренной зоны, насчитывают сходные количества таксонов. Так, на Камчатке с прилегающими островами известно 227 видов печеночников, на о-ве Хоккайдо - 182, в Корее - 223. Таким образом, число в 200-250 видов является характерным для региональных горных приокеанических флор внетропических областей Западной Пацифики.
При выборе регионов и районов, предпочтение отдано территориям с горным рельефом, поскольку не имело смысла проводить сравнение с равнинными областями Голарктики, насчитывающими ничтожное, по сравнению с Камчаткой, число видов. Большинство территорий расположено в пределах Циркумбореальной флористической области (Тахтаджян, 1978), от Арктики (Чукотка) до неморальных областей (Овернь в восточной Франции), в Восточноазиатской области расположены о-в Хоккайдо (в том числе горы Hidaka, о-ва Rishiri и Rebun), массив Chichibu-Okutama на о-ве Хонсю, п-ов Корея (Korea), юг Приморского края (Южное Приморье), о-ва Итуруп и Кунашир, Южный Сахалин, северо-восточный Китай (г. Chokai, г. Hakusan, г. Hwangshan, район Heilongjiang); в области Скалистых гор располагается штат Айдахо (Idaho); наконец часть провинции Онтарио (Ontario) лежит в Атлантическо-Североамериканской области.
В результате проведенных исследований на Камчатке и прилегающих островах было выявлено 227 видов печеночников: это количество является наивысшим показателем богатства среди региональных флор России, опережая на 35 видов ближайшую к ней по богатству флору огромного региона - республики Якутия. Такое особое положение Камчатки объясняется двумя группами причин объективного и субъективного характера. К первым относится приокеаническое положение региона и горный рельеф и, как следствие, значительное разнообразие местообитаний. Ко вторым - лучшая степень изученности, по сравнению с большинством регионов в азиатской России.
Рассматривая камчатское разнообразие внутри Циркумбореальной флористической области, можно заметить, что ее показатель богатства сходен с другими приокеаническими регионами, но несколько уступает прекрасно изученным и расположенным, целиком или частично, в более теплом климате странам Западной Европы, как Швеция (261 вид), Норвегия (279), Великобритания (291) (Soderstrom et al., 2002) и др.
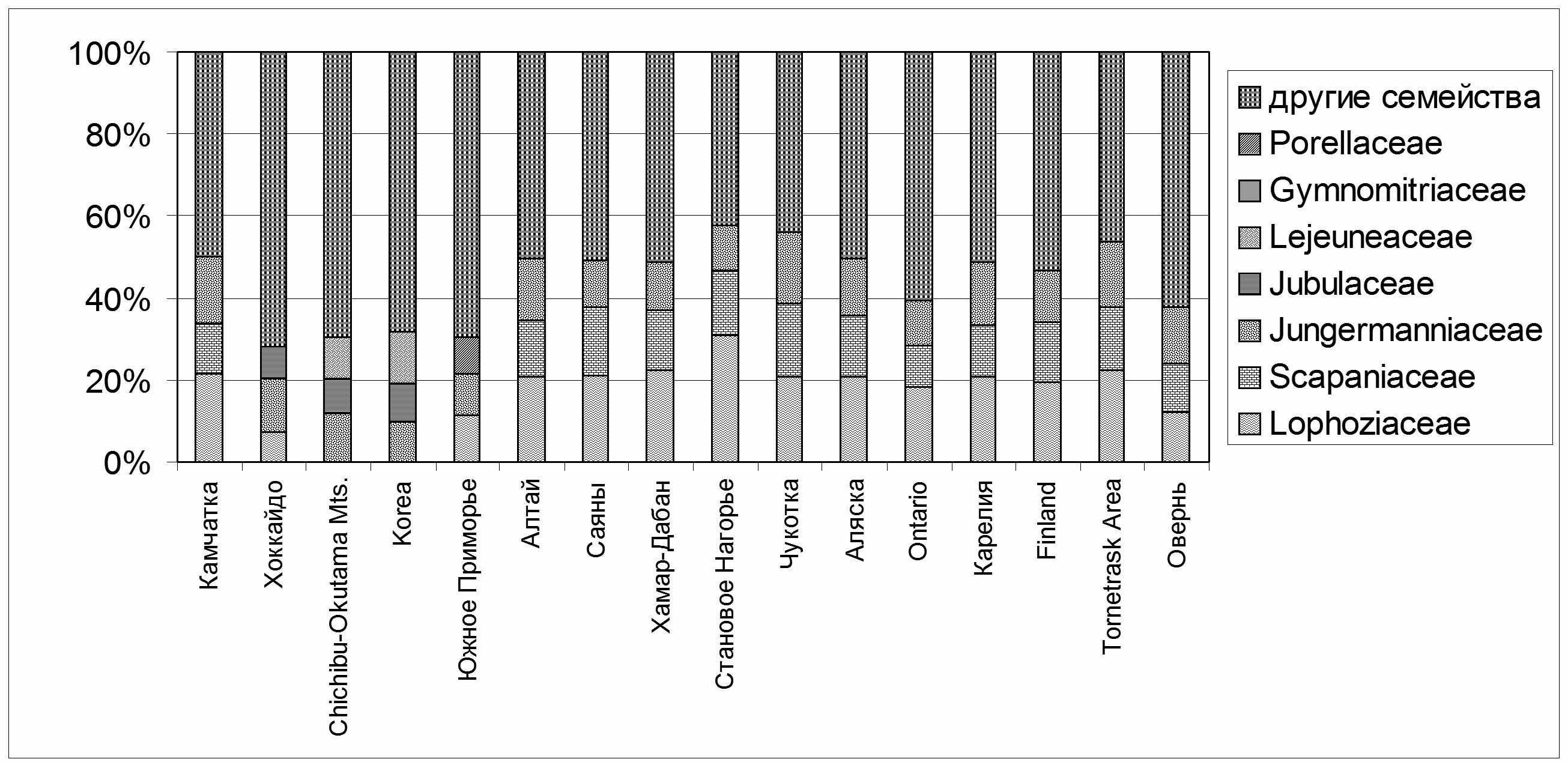 |
| Рис. 3. Вклад ведущих семейств в формирование флор регионов. |
1). Ведущие семейства во флорах севера Голарктики (т.е. расположенных в пределах гипоарктического ботанико-географического пояса (Юрцев, 1966)), включают около 50% видов и более, что полностью согласуется с исследованиями проведенными Константиновой (1998), но южнее, в Онтарио и Оверни, доля ведущих семейств в их флоре составляет уже около 40%. В районах и регионах, находящихся на территории Восточноазиатской области, удельная доля трех ведущих семейств уже колеблется около 30% (Рис. 3).
 |
| Рис. 4. Корреляция между таксономическими родовыми спектрами регионов, при нижней границе коэффициента 90%. (Здесь и далее принята следующая система обозначений сходства: 99-100% − сплошная линия, 97-98% – длинный пунктир, 94-96% – короткий пунктир, 91-93% − длинный пунктир с точкой, 88-90% − длинный пунктир с двумя точками, 85-87% − короткий пунктир с точкой, 82-84% − короткий пунктир с двумя точками, 79-81% − точки, 70-78% − точки через два пробела. 60-70 - волнистая линия) |
2). Во всех голарктических флорах Западной Пацифики, расположенных южнее зональной тундры, т.е. южнее Чукотки, прогрессивно повышается значимость Jungermanniaceae, удельный вес которого сначала превышает Scapaniaceae, а затем, приблизительно с 45° с.ш., и Lophoziaceae, а южнее 40° с.ш. только оно одно входит в тройку ведущих семейств. В Корее Jungermanniaceae следует за Lejeuneaceae и опережает Jubulaceae. Во флоре горного массива Chichibu-Okutama (о-в Хонсю) это семейство стоит на первом месте, следом за ним идут Lejeuneaceae и Jubulaceae. В отличие от западной Пацифики, на востоке ее, даже в гиперокеанических флорах (Алеутские острова, п-ов Аляска) Jungermanniaceae быстро теряет свои позиции. Таким образом, в Восточной Азии удельный вес Jungermanniaceae отражает, во-первых, степень океаничности флоры, причем важным показателем в циркумбореальных флорах является превышение Jungermanniaceae над Scapaniaceae. Во-вторых, выход Jungermanniaceae на первое место в спектре ведущих семейств совпадает с переходом Циркумбореальной области в Восточноазиатскую. Последнее действительно только в островных флорах, на континенте влияние этого семейства быстро падает при удалении от побережья.
Анализ на родовом уровне является более тонким инструментом по сравнению с анализом семейственных спектров. С помощью него было оценено положение флоры в целом как в северной Пацифике, так и в Циркумбореальной области.
Можно предположить, что основным фактором, определяющим близость региональных флор к Камчатке, является приокеаническое положение (и, следовательно, климат). При нижнем пределе коэффициента корреляции в 90% родственными Камчатке оказываются ближайшая Чукотка, а также приатлантические Финляндия и северная Швеция (Tornetrask Area). В целом, выделяется две клады (Рис. 4), расположенные, по случайности, на разных полюсах диаграммы: это - приокеанические Овернь, Камчатка, Карелия, Финляндия, северная Швеция, Чукотка и более континентальные горные системы - Становое Нагорье, Саяны, Хамар-Дабан, Алтай, связанные с Чукоткой и Аляской (необходимо заметить, что хотя оба последние региона располагаются на берегу океана, но в них значительные по протяженности площади заняты ксерофитной растительностью, развивающейся в условиях почти континентального климата). При нижней границе корреляционного коэффициента в 80% происходит образование более или менее тесных связей между всеми регионами в пределах Циркумбореальной области. Наконец, при 60% пороге коэффициента при упрочении циркумбореальных связей намечаются слабые взаимодействия (в виде второй клады) между регионами Восточноазиатской области.
Проведенный корреляционный анализ по спектрам видов показал, что взаимосвязи между флорами весьма слабы и находятся на границе достоверности результата (около 60%). При сравнении регионов, при нижней границе корреляции в 60% (достигнутый максимум - 65%), Камчатка образует связи со всеми вовлеченными в анализ регионами в пределах Циркумбореальной области, за исключением тех, чьи флоры обогащены неморальными элементами: Онтарио и Овернью.
При сравнении видовых списков флористических районов при тех же границах величины коэффициента корреляции выяснено следующее. Все районные флоры изученного региона образуют тесные перекрестные связи, что свидетельствует о сходстве видового состава и о лежащем в его основе длительном сопряженном флорогенетическом развитии в близких климатических и ландшафтных условиях. Интерес представляет также показанное этим анализом родство флор Алеутских островов (не включая Командоры) и п-ова Аляска, горных районов северной окраины Восточноазиатской области, северо-восточной Азии и Арктической Аляски (Рис. 5).
 |
| Рис. 5. Корреляция между таксономическими видовыми спектрами избранных районов, при нижней границе коэффициента 60%. Здесь и далее названия флористических районов полуострова сокращены следующим образом: СК - Северная, ЗК - Западная, ЦК - Центральная, ВК - Восточная, ЮК - Южная Камчатка. |
В долготном отношении подавляющее число таксонов имеет циркумполярный тип ареала (170, или 73%). Виды с другим типом ареала играют в сложении флоры Камчатки существенно меньшую роль. Вместе с тем, заслуживает внимания присутствие довольно значительного количества приокеанических таксонов: 15 (6%) амфиокеанических + 6 (3%) амфипацифических, а также восточноазиатских - 15, или 6%. Участие в формировании флоры представителей других типов ареала ничтожно мало (азиатский - 3, или 1%, евразиатский - 1, или <1%, азиатско-американский - 10 или 4%, восточноазиатский-американский - 1, или <1%, дизъюнктивный - 3, или 1%, мультиареальный - 7, или 3%). Таким образом, флора может быть охарактеризована в долготном отношении как циркумполярная с участием амфиокеанических и восточноазиатских видов. Последняя черта отражает положение Камчатки на восточной окраине азиатского континента.
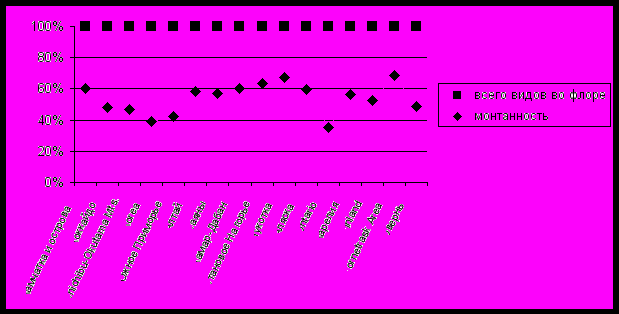 |
| Рис. 6. Относительное количество видов, связанных в распространении с горами в сравниваемых регионах Голарктики. |
Уровень монтанности, широтная и долготная характеристики флоры являются, в данном случае, функцией от климата, вулканической активности и орографии Камчатки и наложенными на эти факторы региональными особенностями флорогенеза. Именно эти факторы обуславливают в каких-то чертах сходство, а в каких-то, наоборот, отличие флоры печеночников Камчатки от других регионов и характеризуют ее флорогенетические связи. Фактически показатель монтанности иллюстрирует генетическую связь флоры с горными системами, а, следовательно, отчасти отражает историю заселения территории.
Как видно из представленной диаграммы (рис. 6), флора печеночников Камчатки и прилегающих островов обладает сходным показателем монтанности с флорами крупных горных систем севера Голарктики - Алтая, Саянами, хр. Хамар-Дабана, Станового Нагорья и регионов с горной орографией - Чукоткой, Аляской, северной Швецией (Tornetrask Area). К группе с более низким показателем по этому параметру относятся либо регионы с преобладающим равнинным или холмистым рельефом (Корея, Приморье, штат Онтарио), либо регионы с богатой пойменной или равнинной растительностью (о-в Хоккайдо, массив Chichibu-Okutama, Овернь). Вместе с тем, заслуживает рассмотрения факт высокой монтанности флор Карелии и Финляндии - регионов, не обладающих крупными горными системами. Это можно объяснить комплексностью монтанности, как фактора отражения характера флоры. Во-первых, этот индекс повышается в районах с хорошо развитыми горными системами, во-вторых, его повышение неизбежно происходит при продвижении к северу. Последнее явление имеет двойную природу: 1) оскудение или исчезновение лесных сообществ и, как следствие, снижение общего таксономического богатства; 2) более широкое распространение пространств, занятых тундровой или тундроидной растительностью и, следовательно, широкое распространение и таксономическое разнообразие видов, на протяжении большей части своего ареала, связанных с тундровыми группировками в горах. Последнее обстоятельство особенно показательно у печеночников. Лишь мизерная часть из таксонов, встречающихся в зональных тундрах, не распространена южнее в горах.
Географическая характеристика изученной флоры сравнивалась с другими регионами с помощью корреляционного анализа, в который для комплексности оценки были вовлечены все долготные и широтные группы. Выяснилось, что подобного рода анализ при применении его к печеночникам показывает сходство и различие в географической характеристике флор лишь в самых общих чертах и подходит, скорее, для сравнения флор различных флористических царств, чем регионов в пределах одной области. Только на самых высоких уровнях корреляции оказалось возможным проследить и выявить неуловимые различия в тенденциях и тяготениях географических спектров.
При достоверности корреляции в 97% образуется единый конгломерат региональных флор северной Голарктики (за исключением более равнинного и пограничного с Восточно-североамериканской областью Онтарио и флоры Оверни, обогащенной видами более южного, в частности, присредиземноморского распространения) (рис. 7). Также намечается связь между регионами, относящимися к Восточноазиатской флористической области - массивом Chichibu-Okutama и Кореей. 95% уровень корреляции показывает еще большее упрочение связей между флорами Циркумбореальной области. Наконец, при 70% уровне корреляции образуются связи между восточноазиатскими региональными флорами и между всеми циркумбореальными. Причем, наиболее северные восточноазиатские регионы образуют достоверную (хотя и слабую) связь с регионами северной Голарктики.
Основным результатом проведенного анализа связей флоры Камчатки и прилегающих островов явилось доказательство ее тесных связей с другими Циркумбореальными региональными флорами и его естественное положение в этой флористическое области. При этом выяснилось, что восточноазиатские связи камчатской флоры печеночников, хотя и присутствуют, но весьма и весьма незначительны. Наибольшим родством флора печеночников изученного региона обладает с другими притихоокеанскими горными флорами севера Голарктики, а также с северными приатлантическими флорами. Несомненна также ее связь с флорами крупных горных систем азиатской России (Алтай, Саяны, Становое Нагорье).
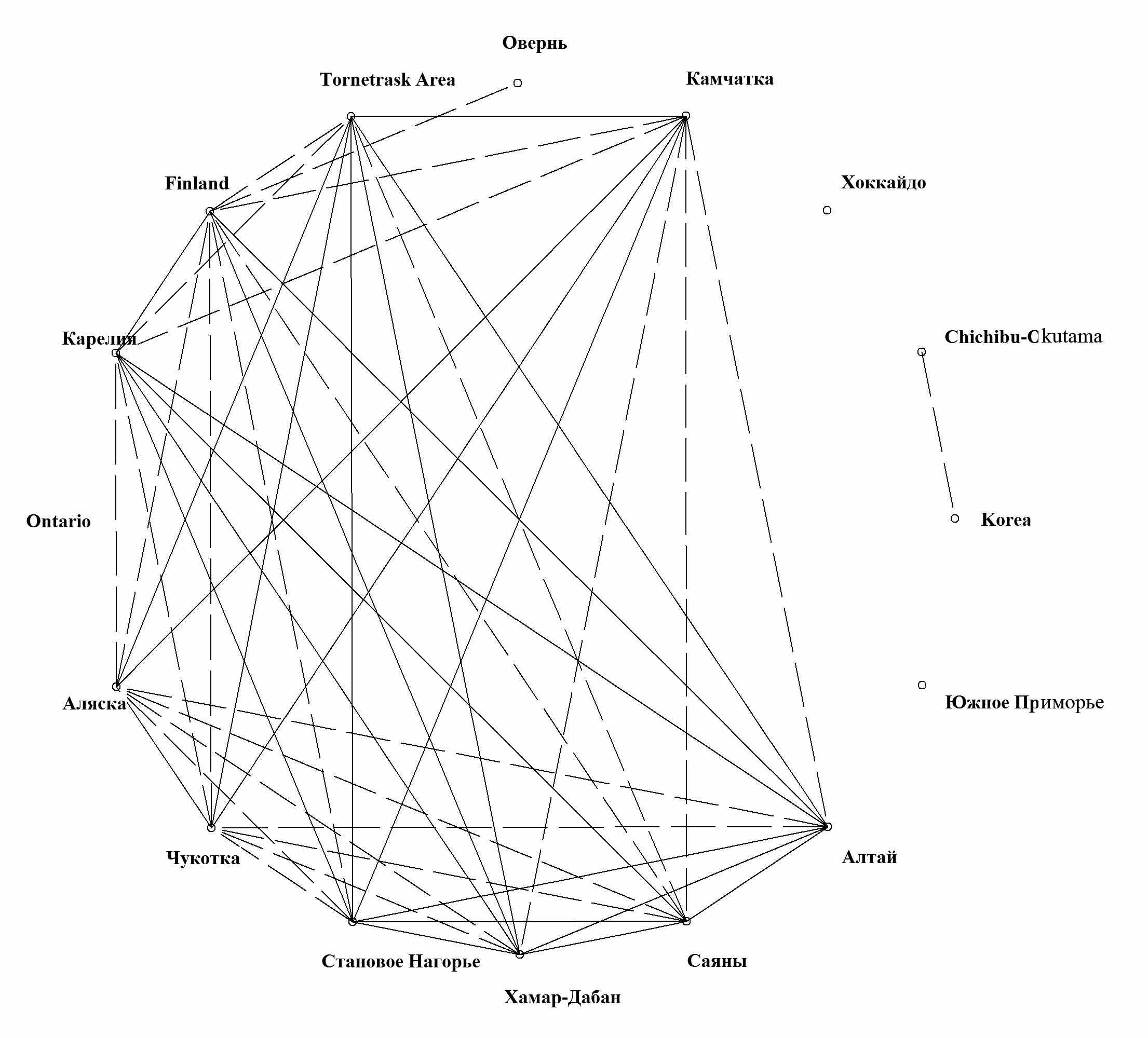 |
| Рис. 7. Корреляция между географическими спектрами флор регионов, при нижней границе коэффициента 97%. |