
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Морфология органов иммуногенеза нерпы байкальской и экспериментальная оценка эффективности ее липидов при разных патологиях
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА2.2. Получение и исследование эффективности липосом
на основе липидов нерпы
Материалом исследований служили липосомы из яичных фосфолипидов; липосомы из фосфолипидов печени нерпы; липосомы из яичных фосфолипидов, загруженные активной фракцией тимуса, отваром противовоспалительного фитосбора; липосомы из фосфолипидов печени нерпы, загруженные водным раствором экстракта какалии копьевидной, водным раствором экстракта черных листьев бадана толстолистного. Все полученные липосомы содержали жир байкальской нерпы в концентрации 3 мг/мл и антиоксидант ?-токоферола ацетат в концентрации 1% (по отношению к липидному компоненту), соответствующий требованиям ФС 42-2654-89-8.10. Жир нерпы получен согласно ТУ 9281-017-02069473-2001.
Экстракцию фосфолипидов из яичных желтков производили по классическому методу Фолча (Биохимия, 1989), из печени нерпы - по методу, предложенному М. Кейтс (1975). Липосомальные структуры готовили по методу, предложенному Bangham A.D. et al. (1964). Ультрамикроскопирование липосомальных частиц производили с помощью электронного просвечивающего микроскопа JEOL JEM-2000 FXII. Количество малонового диальдегида (МДА) как конечного продукта перекисного окисления липидов (ПОЛ) в липосомах определяли методом пробы с 2-тиобарбитуровой кислотой (Сорокоумова Г.М., 2000).
Активную фракцию тимуса получали из вилочковой железы молодняка крупного рогатого скота модифицированным методом В.Я. Ариона (1982). Экстракты какалии копьевидной (ЭКК) (Патент РФ №2206333), черных листьев бадана толстолистного (ЭБТ) (Лубсандоржиева П-Н.Б., 2002), комплексный противовоспалительный фитосбор (Самбатова Э.И., 2001) получены в химико-фармацевтической лаборатории Института общей и экспериментальной биологии СО РАН (ИОЭБ СО РАН). Отвар фитосбора готовили согласно требованиям ГФ XI.
Экспериментальные исследования проводились на: 1) половозрелых мышах обоего пола линии СВА и гибридах (СВАхС57Bl/6) массой 20-22 г, полученных из питомника РАМН Столбовая; 2) белых беспородных крысах обоего пола массой 150-250 г, полученных из питомника филиала №5 ГН - Институт биофизики федерального управления Медбиоэкстрем при Минздраве России г. Ангарск. Животные находились в стандартных условиях содержания в виварии на обычном рационе. Эксперименты осуществляли в соответствии с Правилами проведения работ с использованием экспериментальных животных (Приказ МЗ СССР № 755 от 12.08.77 г.).
Фагоцитарную активность перитонеальных макрофагов (ПМ) мышей в отношении Staph. aureusв системе in vitro исследовали по методике И.С. Фрейдлин (1976). Макрофагальное звено иммунного ответа исследовали в реакциях Fc-розеткообразования и определения антигенпрезентирующей активности макрофагов (Имельбаева Э.А., 1996). Иммуномодулирующую активность липосомальных средств изучали в реакциях, опосредующих клеточный (в реакциях Гиперчувствительность замедленного типа - ГЗТ (Методические рекомендации, 1992) и лтрансплантат против хозяина - РТПХ (Тессенев В., 1979)) и гуморальный (в реакции антителообразования (Методические рекомендации, 1992)) иммунитет. Ростстимулирующую активность тимоцитов морской свинки оценивали в реакции лактивного розеткообразования с эритроцитами кролика (Nekam K. et al., 1982).
Для гистоморфологического исследования участки слепой кишки подопытных мышей и раневые участки крыс фиксировали в 10%-ом растворе нейтрального формалина, депарафинированные гистологические срезы окрашивали гематоксилином и эозином, пикрофуксином по Ван Гизон (Меркулов Г.А., 1969).
Изучение противовоспалительной активности исследуемых средств проводили на стандартных моделях асептического воспаления, лватной гранулемы согласно Методическим рекомендациям (1983) и формалинового отека лапок крыс (Тринус Ф.П., 1975). Также определение противовоспалительной активности проводили на двух моделях острого перитонита (Александров П.Н. с соавт., 1986; Методические рекомендации Е, 1989). Острый адреналиновый отек у мышей вызывали введением раствора адреналина гидрохлорида подкожно (Томчин А.Б. с соавт., 1997).
Оценку мембраностабилизирующей активности липосомальных средств осуществляли по выраженности степени гемолиза эритроцитов, вызываемого реактивом Фентона (перекисный гемолиз), и добавлением дистиллированной воды (осмотический гемолиз) (Ковалев И. Е. с соавт., 1986).
Степень выраженности окислительных процессов в организме экспериментальных животных определяли по содержанию малонового диальдегида в сыворотке крови (Клебанов Г.И. с соавт., 1988) и печени животных (Стальная И.Д., Гаришвили Т.Г., 1977) в модификации Строева Е.А. и Макаровой В.Г. (1986), а также уровня окисленного и восстановленного глутатиона в сыворотке крови (Anderson M.E., 1989).
С целью оценки состояния антиоксидантной системы у животных в сыворотке крови определяли активность глутатионпероксидазы (по методу Stults F.H. et al., 1977), глутатионредуктазы (по методу Racker E., 1955), каталазы (Королюк М.А. и др., 1988) и супероксиддисмутазы (Чевари С. и др., 1985).
Уровни содержания в крови животных липидов (общего холестерина, холестерина липопротеидов низкой и очень низкой плотности, холестерина липопротеидов высокой плотности, триглицеридов) определяли стандартизированными биохимическими методами с помощью наборов реагентов производства ЗАО Вектор-Бест: Новохол, ЛВП-Холестерин-Ново и Триглицериды-Ново.
Гепатопротекторное действие липосомальных средств было изучено на животных c токсическим гепатитом, который вызывали пероральным введением тетрахлорметана согласно (Олейник А.Н., 1984). Желчь для исследования получали в условиях острых опытов по методике Н.П. Скакуна и А.Н. Олейника (1967). Оценку желчеобразовательной и желчевыделительной функции печени проводили по скорости секреции и общему количеству выделенной желчи, концентрации желчных кислот (Карбач Я.И., 1961), а концентрацию холестерина в желчи определяли по методу С.М. Дроговоз (1971), билирубина - по методу Ван ден Берга в модификации Н.П. Скакуна (1956).
Для воспроизведения раневых повреждений были использованы модели линейной, плоскостной кожно-мышечной кожной раны и модель химического ожога кожи (Клюев М.А., 1989; Пономарева-Астраханцева Л.З., 1954; Ойвин И.А., Шетель С.Л., 1961). Прочность образовавшегося рубца определяли методом тензиометрии (Коваленко И.В., 1984); проводили планиметрическое, цитологическое и гистологическое исследования раны (Шалимов С.А., 1989).
Реакции поверхностной иммунофлуоресценции выполняли на клетках, прикрепленных к стеклу с помощью поли-L-лизина (Иммунологические методы, 1987).
Об адаптогенных свойствах липосомальных средств судили по времени выносливости крыс в стандартном тесте плавания до полного утомления (Dawson C.A., Horvath S.M., 1970), по ориентировочно-исследовательскому поведению и суммарной двигательной активности животных в тестах лоткрытое поле (Калуев А.В., 1998) и Т-образный лабиринт (Буреш Я. с соавт., 1991).
Для оценки влияния средств на состояние гематологических показателей у животных определяли концентрацию гемоглобина унифицированным гемоглобинцианидным методом (Пупочная В.И., 2003) с помощью набора реагентов Диагем-Т (НПО РЕНАМ) и вычисляли лейкоцитарную формулу крови общепринятым методом (Меньшиков В.В. с соавт., 1987).
Полученные цифровые данные обрабатывали статистически с применением непараметрических (Вилкоксона-Манна-Уитни) (Гублер И.В., 1973) и параметрических (Стьюдента - с помощью компьютерной программы Jandel SigmaPlot32) критериев. Рассчитывали достоверность отличий данных в группах. Достоверными считались отличия с вероятностью p<0.05.
3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
3.1. Особенности структурной организации
органов иммуногенеза байкальской нерпы в постнатальном онтогенезе
Проведенные исследования по оценке микроанатомических и морфологических характеристик органов иммунной системы байкальской нерпы позволили выявить некоторые особенности их строения как на макро-, так и на микроанатомическом уровнях.
Так, тимус байкальской нерпы, в отличие от тимуса других млекопитающих, состоит не из двух, а из четырех долей. Две доли грудные, расположены за грудиной и лежат на сердце. Две другие доли - шейные, охватывают трахею.
Микроанатомическое строение тимуса нерпы имеет типичный для млекопитающих вид. К видоспецифическим особенностям микроструктуры можно отнести наличие толстой фиброзной капсулы и интенсивное развитие в строме органа волокнистой соединительной ткани, что, по-видимому, объясняется особенностями биологии развития и среды обитания этого животного. Кроме того, по нашим данным, в мозговом веществе тимуса нерпы наблюдается высокое содержание гранулоцитарных лейкоцитов (нейтрофилов и эозинофилов) - более 2%.
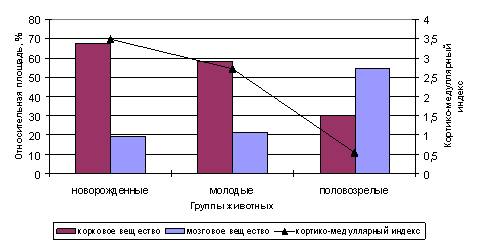 Сравнительный морфометрический анализ показал, что в тимусе новорожденных и молодых нерп преобладает корковое вещество, тогда как у взрослых животных - мозговое. Поэтому кортико-медуллярный индекс, отражающий соотношение площадей коркового и мозгового вещества, с возрастом уменьшается (рис. 3).
Сравнительный морфометрический анализ показал, что в тимусе новорожденных и молодых нерп преобладает корковое вещество, тогда как у взрослых животных - мозговое. Поэтому кортико-медуллярный индекс, отражающий соотношение площадей коркового и мозгового вещества, с возрастом уменьшается (рис. 3).
Рис. 3. Относительная площадь (%) и кортико-медуллярный индекс структурных компонентов тимуса байкальской нерпы в различные возрасты
Исследования показали, что в тимусе половозрелых нерп возрастает доля соединительнотканных элементов и сосудистого русла. Площадь капсулы увеличивается в 1.6 раза, а площадь сосудов - в 3 раза по сравнению с молодыми животными. У взрослых нерп также увеличивается площадь, занимаемая на гистологических срезах тимическими тельцами (в 1.8 раза). Причем, как показали исследования, происходит это не за счет увеличения их количества, а за счет их укрупнения и появления кистоподобных структур. Тем самым, с возрастом у нерп появляются признаки возрастной инволюции.
Исследования морфологии и микротопографии селезенки байкальской нерпы показали, что орган имеет типичную для млекопитающих структурную организацию. Появление в селезенке молодых животных большего, чем у новорожденных, числа лимфоидных узелков с центрами размножения связано с созреванием лимфоидной ткани, усилением ее функциональной активности. Появление высокого содержания плазматических клеток, особенно зрелых (антителопродуцирующих) форм, указывает на резкое усиление гуморального иммунитета с возрастом. При этом слабая морфологическая выраженность зон накопления Т-клеток в периартериальных лимфоидных муфтах (ПАЛМ) связана, видимо, с компенсаторным усилением и преобладающей функцией гуморального иммунитета у этих животных, что объясняется, по-видимому, условиями обитания животных.
Выявлено, что у половозрелых животных, по сравнению с молодыми, четко выявляются признаки возрастной инволюции лимфоидной ткани и органа в целом: резко увеличивается содержание соединительной ткани в селезенке и склерозируются сосуды. В сравнении с молодыми, у половозрелых нерп заметно увеличивается (на 14.8 клетки) плотность клеток в функционально активных лимфоидных узелках с центрами размножения (в центрах размножения и в мантии) (рис. 4), что в значительной степени связано с изменением в соотношении лимфоидных клеток в сторону общего снижения процесса лимфоцитопоэза.
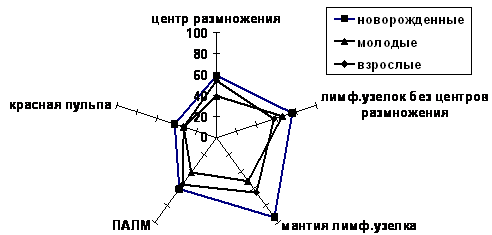
Рис. 4. Соотношение плотности распределения клеток на единице площади гистологического среза (880 мкм2) в структурных зонах селезенки нерп в различные возрастные периоды
У половозрелых нерп, по сравнению с молодыми, в лимфоидных структурах селезенки резко уменьшается количество (вдвое - в центрах размножения) или исчезают (в ПАЛМ и красной пульпе) малодифференцированные формы лимфоцитов. Исключение составляет красная пульпа, где отмечено усиление пролиферации клеток в 1.8 раза. К половозрелому возрасту, по сравнению с молодыми, отмечается тенденция в увеличении содержания зрелых форм лимфоцитов в белой пульпе. При этом у молодых и половозрелых животных практически не изменяется интенсивность процессов деструкции клеток, но несколько снижается макрофагальная активность клеток в центрах размножения и в мантии узелков.
Изучение структурной организации брыжеечных лимфатических узлов байкальской нерпы, принимающих лимфу от тонкого отдела кишечника, выявило следующие особенности: сильное развитие соединительной ткани; большая площадь, занимаемая лимфоидными узелками, высокое содержание среди них вторичных лимфоидных узелков; большая доля, занимаемая мозговым веществом; наличие широких межклеточных пространств.
Клеточный состав брыжеечных лимфатических узлов байкальской нерпы, принимающих лимфу от тонкого отдела кишечника, во всех возрастных группах характеризуется наличием большого числа малодифференцированных форм клеток (лимфобластов и больших лимфоцитов) в центрах размножения лимфоидных узелков; исключительно высоким содержанием плазматических клеток; превышением количества плазмобластов над количеством плазмоцитов; превышением числа малых лимфоцитов над числом средних лимфоцитов почти во всех структурно-функциональных зонах; преобладанием средних и малых лимфоцитов в корковом веществе, по сравнению с мозговым; высоким содержанием ретикулярных клеток; наличием небольшого числа эозинофилов; более высокой плотностью распределения клеток в корковом веществе, по сравнению с мозговым веществом.
В постнатальном онтогенезе брыжеечные лимфатические узлы байкальской нерпы, принимающие лимфу от тонкого отдела кишечника, претерпевают следующие возрастные изменения структурной организации: увеличение относительной площади соединительнотканного компонента; уменьшение доли, занимаемой корковым веществом; уменьшение относительной площади лимфоидных узелков, в том числе вторичных лимфоидных узелков. Проявляется тенденция к возрастному снижению корково-мозгового индекса (рис. 5).
В брыжеечных лимфатических узлах байкальской нерпы, принимающих лимфу от тонкого кишечника, в процессе постнатального онтогенеза происходит снижение числа средних лимфоцитов и ретикулярных клеток, увеличение числа плазматических клеток; уменьшение числа больших лимфоцитов, увеличение содержания малых лимфоцитов в корковом веществе; снижение числа деструктивно измененных и разрушенных клеток в центрах размножения лимфоидных узелков; уменьшение числа малых лимфоцитов в мозговом веществе.
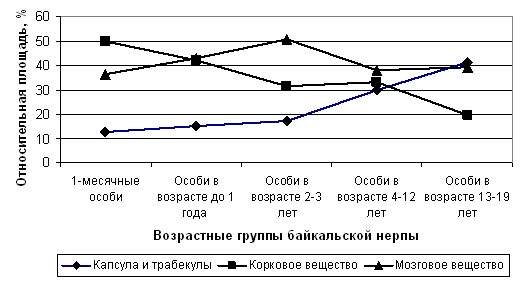
Рис. 5. Динамика возрастных изменений относительной площади основных морфо-функциональных зон брыжеечных лимфатических узлов, принимающих лимфу от тонкого кишечника байкальской нерпы.
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии