
Тема 7 Физиология синапса Синапс
| Вид материала | Документы |
- Физиология памяти и мнемотехника, 242.34kb.
- Методические рекомендации для самостоятельной работы тема, 673.7kb.
- Методические рекомендации для практических занятий тема, 567.31kb.
- Рабочая программа дисциплины «физиология» (физиология растений) Код дисциплины по учебному, 269.31kb.
- Отделения заочного обучения, 25.05kb.
- Программа вступительных испытаний для специальности магистратуры 1-79 80 29 Патологическая, 348.44kb.
- Юрий Александрович Власов лекция, 104.61kb.
- Тема моего реферата физиология памяти и разнообразные способы быстрого и эффективного, 63.49kb.
- «Физиология растений», 308.24kb.
- Рабочая программа дисциплины «физиология» (Физиология животных), 288kb.
Тема 7
Физиология синапса
Синапс – это специализированное структурное соединение между клетками, обеспечивающее взаимное влияние между ними. Через синапсы передаются возбуждающие и тормозные влияния между двумя возбудимыми клетками, осуществляется трофическое влияние, синапсы играют важную роль в реализации механизмов памяти.
Все синапсы классифицируются по следующим критериям:
- По виду соединяемых клеток:
- межнейронные – локализуются в ЦНС и вегетативных ганглиях;
- нейроэффекторные – соединяют эфферентные нейроны соматической и вегетативной нервной системы с исполнительными клетками;
- нейрорецепторные – осуществляют контакты во вторичных рецепторах между рецепторной клеткой и дендритом афферентного нейрона.
- межнейронные – локализуются в ЦНС и вегетативных ганглиях;
- По эффекту: возбуждающие и тормозящие.
- В зависимости от местоположения в ЦНС: аксосоматические, аксодендритные, аксоаксональные, дендросоматические и дедродендритные.
- По способу передачи сигналов:
- Химические – наиболее распространенные в ЦНС, в которых посредником (медиатором) передачи является химическое вещество. Химические синапсы по природе медиатора делят на холинэргические (медиатор – ацетилхолин), адренэргические ( норадреналин), дофаминэргические (дофамин), ГАМК-эргические (γ-аминомасляная кислота) и т.д.
- Электрические, в которых сигналы передаются электрическим током;
- Смешанные синапсы – электрохимические.
- Химические – наиболее распространенные в ЦНС, в которых посредником (медиатором) передачи является химическое вещество. Химические синапсы по природе медиатора делят на холинэргические (медиатор – ацетилхолин), адренэргические ( норадреналин), дофаминэргические (дофамин), ГАМК-эргические (γ-аминомасляная кислота) и т.д.
Механизм синаптической передачи сигналов.
► Химические синапсы – это преобладающий тип синапсов в мозгу млекопитающих и человека. В химическом синапсе выделяют пресинаптическое окончание, синаптическую щель и постсинаптическую мембрану.
В пресинаптическом окончании находятся синаптические пузырьки – везикулы – диаметром до 200 нм, которые образуются либо в теле нейрона и с помощью аксонного транспорта доставляются в пресинаптическое окончание, либо синтезируются (или ресинтезируются) в самом пресинаптическом окончании. Везикулы содержат медиаторы, необходимые для передачи влияния одной клетки на другую. Для синтеза медиатора нужны ферменты, которые образуются в теле клетки на рибосомах и доставляются в пресинаптическое окончание аксонным транспортом. Кроме везикул с медиатором в пресинаптическом окончании имеются митохондрии, которые обеспечивают энергией процесс синаптической передачи. Эндоплазматическая сеть окончания содержит депонированный Са+. Микротрубочки и микрофиламенты участвуют в передвижении везикул. Пресинаптическое окончание имеет пресинаптическую мембрану. Пресинаптической мембраной называют часть пресинаптического окончания, которая ограничивает синаптическую щель.
Синаптическая щель имеет ширину 20-50 нм. В ней содержится межклеточная жидкость и вещество мукополисахаридной природы в виде тяжей между пре- и постсинаптической мембранами. В синаптической щели также находятся ферменты, которые могут разрушать медиатор.
Постсинаптическая мембрана – утолщенная часть клеточной мембраны иннервируемой клетки, содержащая белковые рецепторы, имеющие ионные каналы и способные связывать молекулы медиатора. Постсинаптическую мембрану нервно-мышечного синапса называют также концевой пластинкой.
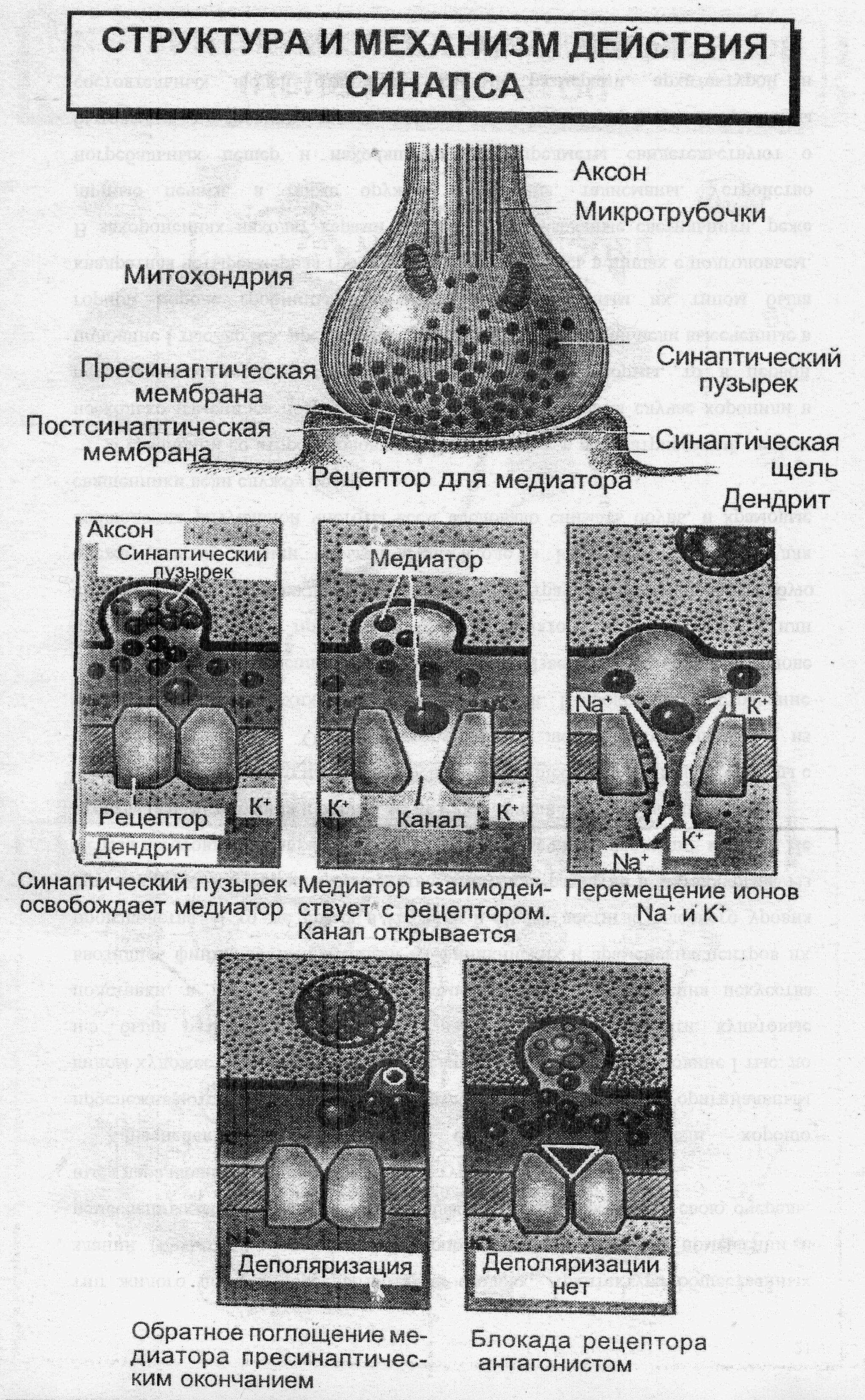
В процессе передачи сигнала в химическом синапсе можно выделить следующие этапы (см. схему):
- Потенциал действия поступает в пресинаптическое окончание.
- После поступления ПД к пресинаптическому окончанию происходит деполяризация мембраны окончания, активируются потенциал-зависимые кальциевые каналы и в синаптическую терминаль входит Са+.
- Повышение концентрации ионов Са+ активирует транспортную систему, что инициирует их экзоцитоз.
- Содержимое везикул выделяется в синаптическую щель.
- Молекулы медиатора, диффундируются в синаптической щели, связываются с рецепторами постсинаптической мембраны.
- Рецепторы постсинаптической мембраны активируют ионные каналы.
- В результате под действием медиатора происходит активация ионных каналов и переход по этим каналам ионов К+ и Nа+ по их градиентам концентрации. Движение ионов формирует постсинаптический потенциал, который по своим свойствам является локальным ответом.
- Медиатор, находящийся в контакте с рецепторами постсинаптической мембраны и в синаптической щели, разрушается ферментами.
- Продукты разрушения медиатора и не разрушенный медиатор всасываются преимущественно в пресинаптическое окончание, где осуществляется ресинтез медиатора и помещение его в везикулы.
На все эти процессы требуется определенное время, которое получило название синаптической задержки и составляет 0,2-0,5 мс. Синаптическая задержка пропорционально зависит от температуры.
Выделение молекул медиатора из пресинаптического окончания пропорционально количеству поступившего туда Са+ в степени n = 4. Следовательно, химическое звено пресинаптического окончания работает как усилитель электрических сигналов.
Химическая передача осуществляет как возбуждающее, так и тормозное действие на постсинаптическую мембрану и зависит от медиатора и рецепторов постсинаптической мембраны. Из пресинаптического окончания выделяются следующие медиаторы:
- Ацетилхолин – встречается в различных отделах ЦНС (кора большого мозга, ретикулярная формация ствола мозга, гипоталамус, спинной мозг). Известен в основном как возбуждающий медиатор (например, он является медиатором ά-мотонейронов спинного мозга). Тормозное влияние ацетилхолин оказывает в глубоких слоях коры большого мозга, стволе мозга и в хвостатом ядре.
- Катехоламины (норадреналин, дофамин, серотонин, гистамин) в основном содержатся в нейронах ствола мозга, в меньших количествах в других отделах ЦНС. Например, амины обеспечивают возникновение процессов возбуждения и торможения в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
- Норадреналин. Норадренэргические нейроны сконцентрированы в основном в области голубого пятна (средний мозг). Норадреналин является тормозным медиатором клеток Пуркинье мозжечка и возбуждающим – в гипоталамусе, ядрах эпиталамуса. В ретикулярной формации ствола мозга и в гипоталамусе обнаружены ά – и β – адренорецепторы. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений.
- Дофамин. Дофаминэргические нейроны имеются в составе полосатого тела, в гипофизе. Дофамин участвует в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения.
- Серотонин. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. С помощью серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре – тормозящие. Серотонин ускоряет процессы обучения, формирование болевых ощущений, сенсорное восприятие, засыпание.
- Гистамин в довольно высокой концентрации обнаруживается в гипофизе и срединном возвышении гипоталамуса. В остальных отделах содержание гистамина очень низко.
- Норадреналин. Норадренэргические нейроны сконцентрированы в основном в области голубого пятна (средний мозг). Норадреналин является тормозным медиатором клеток Пуркинье мозжечка и возбуждающим – в гипоталамусе, ядрах эпиталамуса. В ретикулярной формации ствола мозга и в гипоталамусе обнаружены ά – и β – адренорецепторы. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений.
- Аминокислоты. Кислые аминокислоты (глицин, γ – аминомасляная кислота – ГАМК) являются тормозными медиаторами в синапсах ЦНС. Глицин работает в спинном мозге, ГАМК – в коре больших полушарий, мозжечке, стволе мозга, спинном мозге. Нейтральные аминокислоты (ά – глутамат, ά – аспартат) передают возбуждение: глутаминовая кислота является основным возбуждающим медиатором. Рецепторы глутамата и аспарагиновой кислоты имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга.
- Полипептиды. К ним относят энкефалины, эндорфины, ангиотензин, люлиберин, олигопептиды, субстанцию Р и пептид, вызывающий δ-сон.
- Энкефалины и эндорфины – медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы, черной субстанции, ядрах промежуточного мозга и голубого пятна спинного мозга. Энкефалины и эндорфины дают антиболевые реакции, повышение устойчивости к стрессу и сон.
- Пептид, вызывающий δ-сон также дает антиболевые реакции, повышение устойчивости к стрессу и сон.
- Ангиотензин участвует в передаче информации о потребности организма в воде, повышает артериальное давление, тормозит синтез катехоламинов, стимулирует секрецию гормонов, информирует ЦНС об осмотическом давлении крови.
- Люлиберин участвует в передаче информации о потребности организма в половой активности.
- Олигопептиды – медиаторы настроения, полового поведения, передачи ноцицептивного возбуждения от периферии в ЦНС, формирования болевых ощущений.
- Субстанция Р – является медиатором нейронов, передающих болевую информацию. Особенно много этого полипептида содержится в дорсальных корешках спинного мозга.
- Энкефалины и эндорфины – медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы, черной субстанции, ядрах промежуточного мозга и голубого пятна спинного мозга. Энкефалины и эндорфины дают антиболевые реакции, повышение устойчивости к стрессу и сон.
Кроме выше перечисленных медиаторов, существуют химические вещества, циркулирующие в крови, которые оказывают модулирующее действие на активность синапсов. К ним относятся простагландины и нейрогормоны. Простагландины влияют на секрецию медиатора и работу аденилатциклаз. Гипоталамические гормоны, регулирующие функцию гипофиза, также выполняют медиаторную функцию.
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны и вторых посредников. Например, ацетилхолин в коре большого мозга может вызвать и возбуждение и торможение, в синапсах сердца – торможение, в синапсах гладкой мускулатуры желудочно-кишечного тракта – возбуждение. Катехоламины стимулируют сердечную деятельность, но тормозят сокращения желудка и кишечника.
Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс. Если учащать подачу импульса по аксону, то на каждый последующий потенциал действия ответ постсинаптической мембраны, выраженный величиной изменения трансмембранного потенциала, будет возрастать. Это явление облегчает передачу сигнала в синапсе, усиливая ответ постсинаптического нейрона на очередной раздражитель. Подобное явление получило название «облегчение» или «потенциация». В основе этого процесса лежит накопление кальция внутри пресинаптического окончания при достаточно интенсивной стимуляции. Увеличение количества ионов кальция вызывает экзоцитоз большого количества везикул и, следовательно, большого количества медиатора. Таким образом, большее количество рецепторов на постсинаптической мембране будет активировано и откроется больше ионных каналов, что приведет к большому изменению трансмембранного потенциала на постсинаптической мембране.
Если частота возбуждения пресинаптического окончания в течение короткого времени окажется значительной, то в течение 2-5 минут после ее окончания в ответ на одиночный разряд наблюдается рост амплитуды изменения трансмембранного потенциала в постсинаптической элементе. Механизм этого процесса связывают не только с накоплением ионов кальция в пресинаптической мембране, но и с фосфорилированием белков. Подобный процесс получил название посттетаническая потенциация.
В том случае, когда изменения сохраняются не несколько минут, а в течение десятков минут или даже дней, то говорят о долговременной потенциации. В подобном процессе участвуют сложные метаболические механизмы.
При чрезмерном раздражении пресинаптического окончания в нем происходит истощение медиатора, что приводит к уменьшению амплитуды трансмембранного потенциала на постсинаптической мембране. Этот процесс называется кратковременная депрессия. Он развивается и взаимодействует с посттетанической потенциацией.
В том случае если стимуляция синапса низкочастотна, то возникает десинхронизация по времени активации пре- и постсинаптической мембраны, что приводит в свою очередь к долговременной депрессии данного синапса.
Различная интенсивность использования синапса приводит к его модификации, вызывая улучшение или ухудшение передачи через него сигнала. Синаптическая пластичность имеет большое значение в процессах обучения, забывания, памяти, условных рефлексах.
На постсинаптической мембране под действием медиатора может происходить два основных процесса, связанных с возбуждением и торможением. Электрофизиологическим субстратом этих процессов являются изменения трансмембранного потенциала постсинаптической мембраны, получившие названия возбуждающего постсинаптического потенциала (ВПСП) и тормозного постсинаптического потенциала (ТПСП).
Возбуждающий постсинаптический потенциал. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов натрия. По градиенту концентрации Na+ входят в клетку, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила название: возбуждающий постсинаптический потенциал – ВПСП. ВПСП относится к локальным ответам и, следовательно, обладает способностью к суммации. Выделяют временную и пространственную суммацию.
Принцип временной суммации заключается в том, что импульсы поступают к пресинаптическому окончанию с периодом меньшим, чем период ВПСП. Как следствие, новые порции медиатора выделяются в тот момент, когда трансмембранный потенциал еще не вернулся к уровню мембранного потенциала покоя (МПП). Далее новая деполяризация развивается не с уровня МПП, а с текущего уровня трансмембранного потенциала, который ближе к критическому уровню деполяризации (КУД).
Сущность пространственной суммации заключается в одновременной стимуляции постсинаптической мембраны синапсами, расположенными близко друг от друга. В этом случае ВПСП каждого синапса суммируются.
Если величина ВПСП достаточно велика и достигает критического уровня деполяризации (КУД), то генерируется ПД. Однако не все участки мембраны обладают одинаковой способностью к генерации ВПСП. Так, аксонный холмик, являющийся начальным сегментом аксона относительно сомы, имеет приблизительно в 3 раза более низкий порог электрического раздражения. Следовательно, синапсы, расположенные на аксональном холмике, обладают большей возможностью к генерации ПД, чем синапсы дендритов и сомы. От аксонального холмика ПД распространяется в аксон, а также ретроградно в сому.
Тормозной постсинаптический потенциал (ТПСП). Задачей постсинаптического торможения является снижение возбудимости мембраны нейрона, которое достигается применением тормозящих медиаторов. Например, ГАМК или глицина. Первая, взаимодействуя с рецептором, открывает в постсинаптической мембране хлорные каналы. Это приводит к движению Cl- по электрохимическому градиенту. В результате развивается гиперполяризация, в реализации которой принимают участие и выходящие из клетки К+. В результате гиперполяризации увеличивается расстояние до КУДа и, следовательно, уменьшается возбудимость.
Свойства химического синапса.
- Одностороннее проведение возбуждения, которое осуществляется всегда в направлении от пресинаптического окончания в сторону постсинаптической мембраны.
- замедленное проведение сигнала объясняется синаптической задержкой: необходимо время для выделения медиатораиз пресинаптического окончания, диффузии его к постсинаптической мембране, возникновения постсинаптического потенциала.
- Низкая лабильность синапсов объясняется наличием синаптической задержки и обеспечивает трансформацию ритма возбуждения пресинаптической терминали в ритм возбуждения постсинаптической терминали.
- Проводимость химических синапсов сильно изменяется под влиянием биологически активных веществ, лекарственных средств и ядов, гипоксии.
► Электрические синапсы широко распространены в нервной системе беспозвоночных и низших позвоночных животных. У млекопитающих они имеются в стволе мозга в ядрах тройничного нерва, в вестибулярных ядрах Дейтериса и в нижней оливе. В электрических синапсах узкие щелевые контакты отличаются низким электрическим сопротивлением, в них почти нет токов утечки через внеклеточную среду, поэтому изменения потенциала в пресинаптической мембране могут эффективно передаваться на электрочувствительную постсинаптическую мембрану, которая под воздействием потенциалов действия пресинаптической мембраны изменяет ионную проницаемость и может генерировать потенциалы действия. В электрических синапсах проведение возбуждения происходит без синаптической задержки, ток возможен в обоих направлениях, но легче в одном. Эти синапсы дают возможность получать постоянные, повторяющиеся реакции и синхронизировать активность многих нейронов.