
Социальное поведение и его «энергетическая стоимость» при активации иммунной защиты у лабораторных мышей
| Вид материала | Автореферат диссертации |
СодержаниеМатериалы и методы Результаты исследований Публикации по теме диссертации Акулов А.Е. |
- Герпетические инфекции у человека настоящее и будущее, 458.05kb.
- Урок 28 «Война лягушек и мышей», 1155.42kb.
- Примерная программа дисциплины иммунология. Клиническая иммунология для студентов обучающихся, 327.77kb.
- Тема: органы кроветворения и иммунной защиты, 125.9kb.
- Устройство микропроцессорной защиты для сетей 35-110 кв рзл-02 со свободно программируемой, 117.77kb.
- Центральный выставочный комплекс «Экспоцентр» коротко о выставке название: нефтегаз-2010, 255.27kb.
- Мероприятие 6-57 "Реализация образовательных программ направленных на подготовку молодежи, 218.52kb.
- Агрессивное поведение и пути его коррекции, 103.99kb.
- Налог на добавленную стоимость: его сущность, регулирующая и фискальная роль, 28.79kb.
- Социальное поведение животных, 1088.31kb.
УДК 591.1
На правах рукописи
Акулов Андрей Евгеньевич
СОЦИАЛЬНОЕ ПОВЕДЕНИЕ И ЕГО «ЭНЕРГЕТИЧЕСКАЯ СТОИМОСТЬ» ПРИ АКТИВАЦИИ ИММУННОЙ ЗАЩИТЫ У ЛАБОРАТОРНЫХ МЫШЕЙ
Специальность 03.00.13 - физиология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
НОВОСИБИРСК - 2009
Работа выполнена в лаборатории физиологических адаптаций позвоночных животных Института систематики и экологии животных Сибирского отделения Российской Академии Наук
Научный руководитель:
доктор биологических наук,
профессор М. П. Мошкин
Официальные оппоненты:
доктор биологических наук А.В. Куликов
доктор медицинских наук Е.Л. Альперина
Ведущее учреждение: Институт проблем экологии
и эволюции им. А.Н. Северцова РАН
Защита состоится ______________________ на заседании диссертационного совета Д 001.014.01 при НИИ физиологии СО РАМН (630017, г. Новосибирск, ул. Тимакова, 4)
С диссертацией можно ознакомиться в библиотеке НИИ физиологии СО РАМН
Автореферат разослан ____________________________ 2009 г.
Ученый секретарь диссертационного совета
кандидат биологических наук Бузуева И.И.
Актуальность проблемы. Способность паразитов модифицировать поведение животных широко обсуждается в рамках гипотезы о паразитарной манипуляции хозяином, облегчающей экспансию возбудителей болезней (Poulin, 2000; Thomas et al., 2005), и гипотезы о поведенческих барьерах, выполняющих прямо противоположную функцию (Loehle, 1995; Moshkin et al., 2000). Эффективность распространения паразитов среди хозяев во многом зависит от социального поведения зараженных особей, в частности, от их агрессивности, рост которой повышает вероятность обмена возбудителями (Klein et al., 2004) и которая при инфицировании может, как подавляться (Freeland, 1981; Arnott et al., 1990), так и возрастать (Klein et al., 2004).
В отдельных случаях поведенческие эффекты, наблюдаемые при заражении, обусловлены непосредственным влиянием паразитов на системы нейроэндокринной регуляции хозяев (Мошкин и др., 2003; Thompson, 1983). Однако, более общий механизм паразитарной модификации поведения следует искать в тех нейроэндокринных событиях, которые развиваются вслед за активацией чужеродными антигенами защитных систем хозяина (Мошкин и др., 2003). Несмотря на значительное число работ, посвященных изменению нейроэндокринной регуляции при активации различных звеньев иммунной системы (Корнева, Шхинек, 1988; Besedovsky, Sorkin, 1975; Blalock, 1994; Dantzer, 2001; Devoino et al., 2003; Idova et al., 2008), поведенческие и хемокоммуникационные последствия такой активации изучены в гораздо меньшей степени. В частности, практически не исследовано влияние антигенной стимуляции специфического иммунитета на социальное поведение самцов.
Представление о компромиссном распределении ресурсов организма между конкурирующими функциями (trade–off) предполагает реципрокные отношения между затратами на защиту от инфекции и на морфологические и поведенческие половые демонстрации, включая прямую и сигнальную конкуренцию между самцами (Мошкин и др., 2003; Folstad, Karter, 1992; Wedekind, Folstad, 1994). Но эта конкуренция будет иметь реальное значение только в том случае, если для активации иммунной системы требуются значительные энергетические или пластические ресурсы. Вместе с тем, прямые оценки энергетической стоимости иммунной защиты дают противоречивые результаты. Работы по изучению энергетических затрат на обеспечение специфического иммунного ответа, выполненные на птицах, свидетельствуют, как о статистически достоверном повышении энергообмена во время специфического антителообразования (Ots et al., 2001; Martin II et al., 2003), так и об отсутствии такового (Svensson et al., 1998; Mendes et al., 2006; Nilsson et al., 2007). Оценка энергетической стоимости неспецифической иммунной защиты показывает, что активация иммунитета часто сопровождается снижением спонтанной активности животных и, как следствие, падением энергообмена (Кондратюк, 2005). Однако, затраты энергии в пересчете на условную единицу активности оказываются существенно выше по сравнению с контролем. Следует отметить, что корректное измерение КПД мышечной деятельности, основанное на регистрации спонтанной активности и энергообмена животных, является едва ли выполнимой задачей. Это обусловлено, прежде всего, методологическими трудностями расчета механической работы, производимой животным при реализации разных форм поведения. Поэтому для оценки с эколого-эволюционных позиций энергетической значимости иммунной защиты представляется оправданным поиск ответа на следующий вопрос – влияет ли активация неспецифической или специфической иммунной системы на энергетические затраты, связанные с решением конкретной этологической задачи, например, с установлением социальной иерархии.
Цель работы: изучить влияние активации неспецифического и специфического звеньев иммунной системы на социальную конкурентоспособность и энергетическую стоимость социального поведения у самцов мышей.
Задачи исследования:
1. Исследовать социальную конкурентоспособность, энергообмен и устойчивость приобретённого социального статуса у животных с активированной неспецифической линией иммунной защиты;
2. Изучить влияние дефицита доступной энергии на способность самцов доминировать в условиях социального конфликта;
3. Исследовать социальную конкурентоспособность и энергообмен самцов мышей в разные сроки с момента активации специфической линии иммунной защиты;
4. Изучить роль хемосигналов в формировании и фиксации отношений доминирования/подчинения при социальном конфликте контрольных и антигенстимулированных самцов.
Положения, выносимые на защиту:
1. Активация неспецифического звена иммунной защиты приводит к снижению конкурентоспособности самцов в тестах попарного ссаживания, но не приводит к изменению социального статуса в уже сложившихся иерархических отношениях;
2. Введение липополисахарида (ЛПС) повышает энергетические затраты на установление социальной иерархии;
3. Активация специфического звена иммунной защиты вызывает временное снижение способности подопытных самцов к доминированию при конкуренции с контрольными особями;
4. Предваряющее ссаживанию предоставление запаха антигенстимулированного оппонента контрольному самцу на начальной стадии антителообразования способно инвертировать отношения доминирования/подчинения, т.е. приводит к более частому доминированию антигенстимулированных самцов;
5. Развитие специфического иммунного ответа не приводит к росту энергетических затрат на установление социальной иерархии.
Научная новизна. В рамках данного исследования впервые было показано, что активация иммунной системы является достаточным условием для снижения конкурентоспособности животных в процессе формирования социальной иерархии. Но, как было установлено в экспериментах с активацией неспецифического иммунитета при уже сложившихся отношениях доминирования – подчинения, введение антигена не отражается на иерархическом статусе особи. Впервые было выявлено, что предварительное знакомство контрольного самца с запахом мочи антигенстимулированного соперника, находящегося на стадии максимального роста титра антител, способно инвертировать отношения доминирования/подчинения. При этом предварительное знакомство с запахом будущего соперника играет решающую роль в закреплении иерархических отношений.
Впервые исследована энергетическая реакция на социальные стимулы и показано, что прирост энергообмена на запах мочи у самцов, во многом зависит от их будущего социального статуса, установленного в тесте попарного ссаживания, а увеличение энергообмена после ссаживания зависит от статуса и иммунологического состояния животного. Так же впервые установлено, что прирост энергообмена в ответ на ссаживание положительно коррелирует с числом актов агонистического поведения только у ЛПС–стимулированных особей и в большей степени это характерно для доминантных самцов.
Научно-практическая значимость. Перспективы изучения механизмов, лежащих в основе паразитарного влияния на поведение хозяина, заключаются в понимании того, как разные пути иммунного реагирования на паразита влияют на разные формы поведения животного. Полученные нами данные вносят вклад в решение фундаментальной проблемы механизмов паразитарного «манипулирования».
Результаты исследования могут быть полезными при разработке зоотехнических технологий, направленных на оптимизацию условий содержания в неволе животных со сложной социальной структурой. Изменения сложных форм поведения и их энергетической стоимости при активации специфического иммунитета расширяют существующие представления о возможных побочных эффектах вакцинации.
Диссертационный материал используется при чтении курсов лекций «Экологическая физиология» и «Популяционная физиология» в Новосибирском и Томском государственных университетах.
Апробация работы. Результаты работы были представлены на российских и зарубежных конференциях: «XLII Международная научная студенческая конференция. Биология» (Новосибирск, НГУ, 2004), «XXXI апрельская конференция студентов и молодых ученых КемГУ» (Кемерово, КемГУ, 2004), «Сибирская зоологическая конференция» (Новосибирск, 2004), «Международная конференция проблемы популяционной экологии животных» (Томск, 2006), «Химическая коммуникация животных. Фундаментальные проблемы» (Москва, 2006), «6th International symposium on Physiology and Behaviour of Wild and Zoo Animals» (Germany, Berlin, 2007), «V European Congress of Mammalogy» (Italy, Siena, 2007).
Материалы диссертации докладывались на отчетной сессии ИСиЭЖ СО РАН (Новосибирск, 2007).
Публикации. По теме диссертации опубликовано 9 работ (2 из них в рецензируемых журналах).
Структура работы. Диссертация состоит из введения, обзора литературы, главы посвященной поведенческим и энергетическим эффектам активации неспецифического звена иммунной защиты, главы посвященной поведенческим и энергетическим эффектам активации специфического звена иммунной защиты, обсуждения, выводов и списка используемой литературы, который включает 220 названий. Работа изложена на 143 страницах. Иллюстративный материал представлен 11 таблицами и 33 рисунками.
МАТЕРИАЛЫ И МЕТОДЫ
Работа выполнена в 2003–2006 годах на лабораторных мышах аутбредной линии ICR, которых получали из вивария Института цитологии и генетики СО РАН и ГНЦ вирусологии и биотехнологии «Вектор». Перед проведением экспериментальных работ животных в течение двух недель передерживали в условиях вивария лаборатории. Животных содержали в индивидуальных пластиковых клетках (30х20х10) в присутствии запаха половозрелых самок, комнатной температуре (22ºС) и искусственном фотопериоде (14 свет : 10 темнота). В качестве подстилки использовали сухие древесные опилки. Корм и гранулированный корм давали ad libitum. Для активации неспецифического звена иммунной системы животным за 3 часа до тестов вводили ЛПС (L–4005, E.coli, серотип 055:B5, Sigma) в дозе 50 мкг/кг. Для активации специфического звена иммунной системы использовали введение эритроцитов барана (ЭБ) (0.5 мл 2% суспензии ЭБ, содержащей 4х108 клеток/мл). Для формирования дефицита доступной энергии использовали 2–диокси–d–глюкозу (2ДГ), которую вводили за 40 минут до тестирования в дозе 500 мкг/кг. Контрольным животным вводили физиологический раствор. Все виды инъекций проводили внутрибрюшинно. Эксперименты выполнены в соответствии с правилами проведения работ на экспериментальных животных (Приложение к Приказу Министерства здравоохранения №267 от 19.06.2003).
Измерение энергообмена и проведение попарного ссаживания производили в термостабилизированной камере с внутренней температурой 25ºС и постоянной циркуляцией атмосферного воздуха. Энергообмен исследовали методом непрямой калориметрии (при помощи газоанализатора ПЭМ–2М измеряли выделенное животными СО2 , разрешающая способность прибора составляла 0.002%).
Для удобства далее в общей схеме тестирования принято следующее сокращение «а» эксперименты с активацией неспецифического звена и «б» эксперименты с активацией специфического звена иммунной системы.
Общая схема тестирования: 1) помещение животных («а» – на 60 минут и «б» – на 30 минут) в индивидуальные респирометрические камеры («а» – объемом 14 х 22 х 9 см и «б» – объемом 200 мл) для регистрации фонового энергообмена, 2) в экспериментах «б» в варианте с предварительным представлением запаха оппонента предтестовая регистрация длилась не 30, а 20 минут + в течение 10 минут представляли запах мочи оппонента в виде 10 мкл мочи нанесенной на фильтровальную бумагу (2 х 7 см), 3) проведения 15–и минутного попарного ссаживания, 4) продолжение регистрации индивидуальных значений энергообмена («а» – в течение 80 минут после ссаживания, и «б» – в течение 30 минут после ссаживания). При этом использовали следующие показатели энергообмена: 1) фоновый энергообмен (среднее значение выделенного животным СО2 за 4 минуты до предъявления социальных стимулов – предоставление запаха или ссаживание), 2) энергетическая реакция на предоставление запахового стимула – суммарное отклонение выделения СО2 от ожидаемого уровня с 3 по 10 минуту после предъявления стимула (только в варианте с предварительной экспозицией запаха в «б»), 3) энергетическая реакция в ответ на ссаживание («а» – значение уровня выделенного СО2 на 6 минуте после проведения ссаживания, «б» – сумма отклонений выделения СО2 от ожидаемого уровня с 4 по 22 минуту после окончания ссаживания).
Поведение в тесте попарного ссаживания записывали через веб–камеру на компьютер в режиме реального времени. При обработке видеозаписей использовали компьютерную программу, которая обеспечивала автоматическое суммирование числа актов поведения (в экспериментах с активацией неспецифического звена иммунной системы) и их общую продолжительность (в экспериментах с активацией специфического звена иммунной системы). Предварительное, за день до тестирования, выстригание небольших участков шерсти на спине у самцов позволяло фиксировать следующие индивидуальные акты социального взаимодействия: 1) неагонистические (вертикальные стойки с исследовательской целью, назо–назальные и назо–генитальные обнюхивания), 2) агонистические, которые можно разделить на агрессивные (атаки, преследования, биения хвостом, укусы) и субмиссивные (защитные стойки, вокализации, выпады, убегания). Для определения социального статуса самца, вычисляли индекс агрессивности (ИА) по формуле: ИА = (агрессивные акты – субмиссивные акты) / (сумма агрессивных и субмиссивных актов). Животных с отрицательным ИА относили к подчиненным, с положительным к доминантам.
Максимальное потребление кислорода (МПК), отражающее функциональные резервы, оценивали методом непрямой калориметрии в установке закрытого типа (Калабухов, 1951; Шилов, 1961). Животных помещали в герметичную камеру объемом 1.5 л, воздух в которой принудительно прокачивался через капсулу с сухой щелочью. Потребление кислорода определяли по падению давления в данной системе. Измеряющий манометр был прокалиброван в единицах объема. Для оценки интенсивности метаболизма в условиях глубокого охлаждения использовали 15 мин. экспозицию при +7 - +8С в гелиево-кислородной (Helox) атмосфере (Rosenmann, Morisson, 1974; Wang, 1980). Сразу после помещения животного в камеру через нее в течение 3-х минут с помощью поршневого насоса прокачивали гелиево-кислородную смесь (80:20), общий объем которой не менее чем в три раза превышал объем камеры. После прекращения подачи газовой смеси, на 1, 3, 5, 8, 10, 13, 15 минутах измеряли минутное потребление кислорода животным. Убыль кислорода в измерительной системе пополняли на 6-7 и 11-12 минутах.
Влияние социального статуса и иммунного состояния на фоновый энергообмен и прирост энергообмена на запах и ссаживание оценивали с помощью многофакторного дисперсионного анализ (ANOVA). При сравнении 2-х средних использовали t-критерий Стьюдента (t) и критерий Манн-Уитни (Z). Для установления зависимости величины прироста энергообмена от количества агонистических актов рассчитывали коэффициенты корреляции Пирсона и линейной регрессии. При ненормальном распределении вариационных рядов, в частности, при анализе закрепления установившихся иерархических отношений в тестах попарного ссаживания рассчитывали коэффициент ранговой корреляции Спирмена для ИА, зарегистрированных при первом и повторных тестированиях.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Влияние стимуляции неспецифической иммунной системы на социальное поведение и энергообмен самцов
Активации неспецифической иммунной защиты организма приводит к появлению у животных характерного комплекса поведенческих реакций, объединяемых понятием «синдром болезненного поведения» (Hart, 1988; Dantzer, 2001), при этом следует ожидать снижение конкурентоспособности таких особей. В наших экспериментах введение ЛПС вызывало отчетливое подавление способности ЛПС–стимулированных самцов становиться доминантами, при условии, что социальное взаимодействие животных происходило впервые. Об этом свидетельствуют низкие значения ИА ЛПС–стимулированных самцов по сравнению с контрольными животными (-0.37 ± 0.18 и 0.37 ± 0.18, соответственно, Z = 2.37, n1 = 8, n2 = 8, p < 0.05). Но если иерархические отношения между особями сложились во время предварительного 15 мин попарного взаимодействия, то введение ЛПС только доминанту не отражалось на его способности доминировать при повторном ссаживании с тем же партнером. Несмотря на общее снижение интенсивности агонистических взаимодействий при повторном ссаживании, получившие ЛПС доминантные особи достоверно превосходили контрольных подчиненных самцов по ИА (0.22 ± 0.14 и -0.22 ± 0.14, соответственно, Z = 1.94, n1 = 8, n2 = 8, p < 0.05).
Наряду с поведенческими эффектами введения ЛПС, активация неспецифического звена иммунной системы отражается и на энергетическом обмене животных. В тестах, поставленных таким образом, что пары состояли либо из контрольных, либо из ЛПС–стимулированных особей, фоновый энергообмен зависел от иммунного статуса (F1,62 = 33.52, p < 0.001) и был ниже у животных, которым вводили ЛПС. Ссаживание приводило к увеличению энергообмена (рис. 1). При этом прирост энергообмена определялся как иммунным состоянием особи (F1,62 = 6.85, p < 0.05), так и его социальным статусом, установленным непосредственно во время конфликта (F1,62 = 6.25, p < 0.05). Доминантные животные, тестированные после введения ЛПС, показывали больший прирост СО2 по сравнению с доминантами, исследованными на фоне физиологического раствора (t = 2.27, n1 = 10, n2 = 7, p < 0.05). У подчиненных самцов, получивших ЛПС, прирост энергообмена не отличался от такового у подчиненных животных, которым вводили физиологический раствор (t = 1.68, n1 = 10, n2 = 7, p > 0.05).
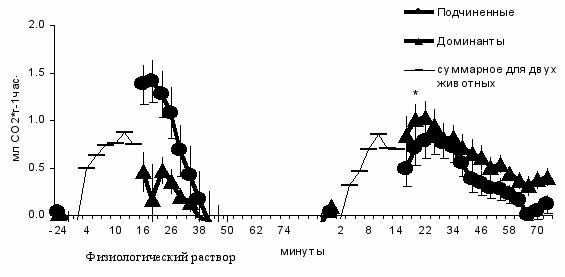
ЛПС
Рис. 1. Динамика энергообмена при 15 мин взаимодействии самцов в тесте попарного ссаживания (выделения СО2 рассчитано на суммарную массу соперников) и в период восстановления (индивидуальные значения выделения СО2).
* - p < 0.01; по сравнению с доминантами, которым вводили физиологический раствор (t-тест Стьюдента).
Стоит отметить, что введение ЛПС приводит как к поведенческим, так и энергетическим изменениям у животных и более того, прирост энергообмена в ответ на ссаживание во многом может определяться поведением животного в ходе этого ссаживания. Так было установлено, что при ссаживании на фоне ЛПС число агонистических актов, проявленных в тесте, коррелирует с приростом энергообмена (рис. 2). Коэффициент линейной регрессии, отражающий угол наклона для зависимости прироста энергообмена от количества агонистических актов у животных, протестированных на фоне введения ЛПС, статистически значимо выше (0.053 ± 0.016), чем у самцов, протестированных на фоне физиологического раствора (0.0032 ± 0.0085, t = 2.77, n1 = 20, n2 = 14, р < 0.05). Примечательно, что для неагонистических актов коэффициенты корреляции с приростом энергообмена были статистически незначимы.
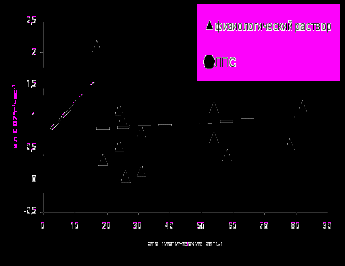
Рис. 2. Зависимость прироста энергообмена (выделение СО2) от количества агонистических актов, проявленных животными в 15 мин тесте попарного ссаживания.
Влияние ограничения доступной энергии на социальную конкурентоспособность самцов
Одной из причин снижения социальной конкурентоспособности при введении ЛПС может быть дефицит доступной энергии, потребность в которой выше при агонистических взаимодействиях у ЛПС-стимулированных доминантов, чем у контрольных. Значимость энергетического фактора для установления социальной иерархии была исследована в экспериментах с введением 2ДГ. Инъекции препарата одному из партнеров за 40 мин до ссаживания не оказывали достоверного влияния на способность к доминированию. Соотношение доминантов и починенных было равно 3/5 в группе 2ДГ и 5/3 в группе их контрольных соперников. ИА у контрольных самцов и самцов, которым вводили 2ДГ, также достоверно не различались (0.25 ± 0.36 и -0.25 ± 0.36, соответственно, Z = 0.97, n1 = 8, n2 = 8, p > 0.05).
Вместе с тем, используемая нами доза 2ДГ значимо влияла на аэробную производительность животных, которую оценивали по максимальному потреблению кислорода в условиях холода (рис. 3). Контрольные и ЛПС–стимулированные самцы имели сходные значения МПК (10.48 ± 1.03 мл*г-1час-1 и 9.64 ± 1.49 мл*г-1час-1, соответственно, t = 0.48, n1 = 8, n2 = 5, p > 0.5). А животные, которым вводили 2ДГ, показывали почти трехкратное снижение максимальной аэробной производительности: МПК = 3.49 ± 0.79 мл*г-1час-1, t = 4.39, p < 0.01 по сравнению с контролем и t = 3.37, p < 0.05 по сравнению с ЛПС–стимулированными самцами.
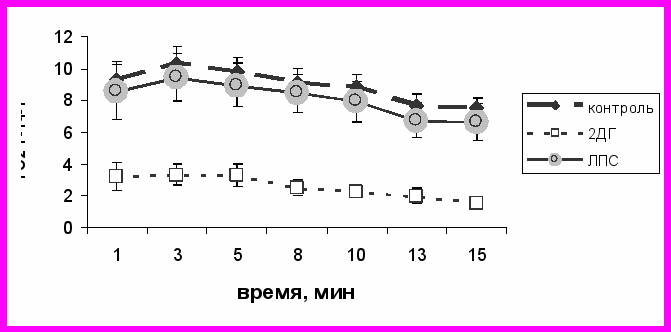
*
Рис. 3. Динамика потребления кислорода животными при экспозиции гелиево-кислородной смесью контрольных самцов и самцов, которым вводили 2ДГ и ЛПС. Среднее ± S.E.
* - p < 0.01 для значений минутного потребления кислорода на протяжении всего теста по сравнению с контрольными и ЛПС–стимулированными самцами (t-тест Стьюдента).
Влияние антигенной стимуляции и запаха мочи оппонента на социальное поведение и энергообмен самцов
Активация специфического звена иммунной защиты у самцов приводит к снижению их конкурентоспособности в тестах, выполненных без предварительного знакомства с запахом соперника через 3 часа и 3 суток после инъекции ЭБ. Через 3 часа после инъекции ЭБ контрольные животные имели более высокий ИА, чем антигенстимулированные (0.54 ± 0.24 и -0.40 ± 0.27, Z = 1.77, n1 = 10, n2 = 10, p < 0.05, соответственно; односторонний критерий Манн–Уитни). Схожий результат был и при ссаживании без предварительного знакомства с запахом соперника на 3 сутки, где ИА у контрольных самцов были выше, чем у животных, которым вводили ЭБ (0.62 ± 0.29 и -0.43 ± 0.36, Z = 1.35, n1 = 5, n2 = 5, p < 0.05). Если в тестах, выполненных через 3 часа возможность предварительного знакомства с запахом оппонента не оказывает существенного влияния на социальное поведение, то в тестах, выполненных на 3–е сутки, это влияние было значимым. Тестирование в этот срок показало, что предварительное знакомство с запахом оппонента приводит к доминированию антигенстимулированных животных. ИА у антигенстимулированных животных был достоверно выше, чем у контрольных особей (0.53 ± 0.19 и -0.46 ± 0.21, Z = 2.48, n1 = 14, n2 = 14, p < 0.05, соответственно).
Инверсия в соотношении доминантов и подчиненных среди контрольных и антигенстимулированных особей сочеталась с различной энергетической реакции на запах будущего соперника у животных разных экспериментальных групп и разного иерархического статуса. При тестировании через 3 дня после введения ЭБ или физиологического раствора было установлено, что среди контрольных самцов особи, ставшие доминантами, характеризуются меньшим приростом энергообмена в ответ на запах соперника, чем особи, ставшие подчиненными. Среди антигенстимулированных самцов это соотношение было прямо противоположным (рис. 4). Сравнение разных форм социального поведения контрольных доминантов в ссаживаниях, где их оппонентом являлся либо контрольный самец, либо антигенстимулированный, не выявило различий по суммарной продолжительности агрессивных актов.
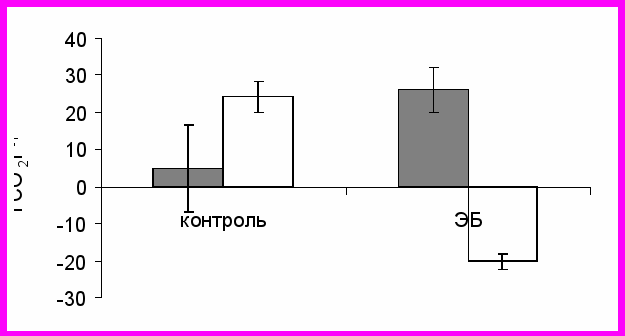
**
*
Рис. 4. Прирост энергообмена (суммарное отклонение выделения СО2 от ожидаемого уровня) в ответ на предоставление запаха оппонента в тестах, выполненных на 3-и сутки после введения физиологического раствора или ЭБ. Среднее ± S.E. Темные столбики – доминантные самцы, светлые столбики – подчиненные животные.
* - p < 0.05; ** – p < 0.01 по сравнению с доминантными самцами.
При этом, доля прямой агрессии (атака + преследование + укус) в общем комплексе агрессивного поведения была меньше у контрольных доминантов в тестах, в которых им оппонировали антигенстимулированные особи (75.3 ± 4.6 %, n = 7), по сравнению с таковой в тестах – «контроль против контроля» (94.2 ± 4.1 %, n = 4, p = 0.02, тест Манн-Уитни). Вместе с тем, при взаимодействии с антигенстимулированными подчиненными контрольные доминанты показывали большую долю биений хвостом, демонстрирующую их агрессивные намерения (24.7 ± 4.6 %, n = 7), чем это наблюдалось при ссаживании двух контрольных самцов (5.8 ± 4.1 %, n = 4, p = 0.02, тест Манн-Уитни). Иными словами, при отсутствии достоверных различий по общей продолжительности агрессии у контрольных доминантов в структуре агрессивного поведения происходили существенные изменения, зависящие от принадлежности подчиненных к контрольным или антигенстимулированным особям. В последнем случае происходило увеличение доли демонстрационной агрессии и снижение доли прямой агрессии.
При первом ссаживании контрольных и антигенстимулированных самцов через 6 суток после инъекции их социальная конкурентоспособность не различалось. ИА был равен 0.19 ± 0.23 у контрольных особей и -0.15 ± 0.24 у антигенстимулированных (Z = 0.84, n1 = 16, n2 = 16, p = 0.38).
Предварительное знакомство с запахом друг друга играло решающую роль в закреплении иерархических отношений, установившихся при первом ссаживании, несмотря на то, что физиологическое состояние и запаховые сигналы антигенстимулированных самцов изменялись в ходе иммунного ответа. Повторные ссаживания, выполненные через 3 и 6 суток после введения ЭБ, показали, что в опытах без предварительной экспозиции запаха или с предварительной экспозицией, иерархические отношения, сложившиеся в первом ссаживании у большинства пар не меняются. О чем свидетельствуют статистически значимые коэффициенты корреляции индексов агрессивности, зарегистрированных через 3 часа и 3-е суток после введения препаратов (r = 0.83, n = 8, p < 0.01 –для тестов без запаха и r = 0.66, n = 12, p = 0.01 – для тестов с предварительной экспозицией запаха мочи оппонента). При ссаживании на 6-е сутки в варианте без предварительной экспозиции запаха было отмечено, что во многих парах доминант становиться подчиненным, а подчиненный доминантом, при этом индексы агрессивности практически не коррелировали (r = 0.03, n = 8, p = 0.93). В ссаживаниях с предварительной экспозицией запаха, иерархические отношения, наблюдаемые в тестах выполненных через 3-е суток, не изменяются ни в одной из пар протестированных на 6-е сутки (r = 0.97, n = 12, p < 0.001).
В эксперименте в котором первое ссаживание происходило через 3 дня после инъекции ЭБ ситуация выглядит похожим образом. Иерархические отношения, сложившиеся через 3 дня в опытах без экспозиции запаха не воспроизводятся в опытах проведенных через 6 дней (r = 0.19, n = 10, p > 0.5), а в варианте с предварительной экспозицией воспроизводятся на статистически значимом уровне (r = 0.62, n = 28, p < 0.001).
Таким образом, предварительная экспозиция запаха играет существенную роль в фиксации ранее установленных иерархических отношений.
Заключение. Полученные нами результаты свидетельствуют о том, что введение бактериального эндотоксина приводит к росту энергетических затрат на установление социальной иерархии. В этом отношении эффект ЛПС, запускающий неспецифический иммунный ответ, принципиально отличается от последствий антигенной стимуляции специфического иммунитета, когда процесс образования специфических антител, индуцированный введением ЭБ, практически не влияет на энергетическую стоимость социального поведения у самцов мышей. Сопоставление эффектов ЛПС и ЭБ, адресованных к разным звеньям иммунной системы, подтверждает предположение Рёберга с соавторами (Råberg et al., 2002) о том, что эволюция иммунитета от врожденного (неспецифического) к приобретенному (специфическому), направлена не только на расширение репертуара иммунной защиты, но и на оптимизацию энергетических затрат, связанных с противостоянием паразитам.
ВЫВОДЫ
1. Самцы с активированной неспецифической линией иммунной защиты при первом взаимодействии проигрывают контрольным животным. Введение ЛПС отдельным особям доминантным или подчиненным в уже сложившихся социальных группах не отражается на их иерархическом статусе.
2. Введение ЛПС приводит к росту энергетических затрат на социальные взаимодействия и, в первую очередь, на агонистические формы поведения. При этом больший рост энерготрат наблюдается у доминантных особей.
3. Введение 2ДГ снижает максимальную аэробную производительность самцов, но дефицит доступной энергии не отражается на их способности доминировать в социальном конфликте.
4. На начальных стадиях специфического иммунного ответа снижается способность подопытных самцов к доминированию при конкуренции с контрольными особями.
5. Предварительное знакомство с запахом мочи соперника, собранной на начальной стадии антителообразования, меняет отношения доминирования/подчинения антигенстимулированных и контрольных животных на прямо противоположные.
6. Среди антигенстимулированных самцов достоверный прирост энергообмена, как индикатор эмоциональной реакции на запах, показывают особи, которые становятся доминантами в тесте попарного ссаживания. Среди контрольных самцов, наоборот, энергетическая реакция на запах выше у будущих подчиненных.
7. Предварительное знакомство с запахом будущего соперника играет решающую роль в закреплении иерархических отношений, установившихся при первом ссаживании.
8. Развитие специфического иммунного ответа на чужеродные антигены не приводит к росту энергетических затрат на установление социальной иерархии.
Публикации по теме диссертации:
- Акулов А.Е. Энергозатраты на установление социальной иерархии при активации иммунной системы // XLII Международная научная студенческая конференция. Биология. – Новосибирск, 2004. – С. 75
- Акулов А.Е. Энергетическая стоимость установления социального иерархического статуса // XXXI апрельская конференция студентов и молодых ученых КемГУ. – Кемерово, 2004. – С. 344–345
- Акулов А.Е., Петровский Д.В. Энергозатраты на установление социальной иерархии в норме и при активации иммунной системы // Сибирская зоологическая конференция. – Новосибирск, 2004. – C. 211
- Акулов А.Е., Петровский Д.В. Изменение агрессивного поведения и его энергетической стоимости при активации специфической иммунной защиты // Международная конференция проблемы популяционной экологии животных. – Томск, 2006. – С. 98 – 99
- Акулов А.Е., Петровский Д.В. Поведенческие и биоэнергетические эффекты, обусловленные разным содержанием низкомолекулярных белков мочи у соперничающих самцов лабораторных мышей // Химическая коммуникация животных. Фундаментальные проблемы. – Москва, 2006. – С. 23
- Akulov A.E., Petrovsky D.V. Direct and scent mediated effect of immunoenhancement on the social behavior of laboratory mice // 6th International symposium on Physiology and Behaviour of Wild and Zoo Animals. – Berlin, 2007. – C. 275
- Akulov A.E. Activation of innate immunity increases energetic cost of aggressive behaviour // V European Congress of Mammalogy. – Siena, 2007. – C. 61
- Акулов А.Е., Петровский Д.В., Мошкин М.П. Антигенная стимуляция как фактор, модулирующий социальное поведение и хемосигналы самцов лабораторных мышей // Журнал высшей нервной деятельности им. Павлова. – Москва, 2009. – Т.59. №3, С.335–343
- Акулов А.Е., Петровский Д.В., Мошкин М.П. Энергетическая стоимость социального поведения мышей при активации неспецифического иммунитета // Журнал общей биологии. – Москва, 2009. – Т.70. №4, С.275–284
Автор выражает благодарность научному руководителю и идейному вдохновителю: Михаилу Павловичу Мошкину, научному консультанту и учителю: Дмитрию Валериановичу Петровскому, сотрудникам лаборатории физиологических адаптаций позвоночных животных за помощь в проведении экспериментов: Кондратюк Е. Ю., Новикову Е. А., Мак В. В., Колосовой И. Е., Литвиновой Е. А., а также первому руководителю профессору Кемеровского Госуниверситета Надежде Алексеевне Литвиновой и декану биологического факультета Кемеровского Госуниверситета Галине Викторовне Ефремовой, за предоставление возможности начала данной работы.
Особую признательность выражаю своей семье и близким людям за поддержку при написании и подготовке к защите диссертации.