
Экологические последствия загрязнения тяжёлыми металлами фитоценозов центральной россии 03. 00. 16 Экология
| Вид материала | Автореферат |
- Особенности загрязнения четвертичных отложений территории г. Москвы тяжелыми металлами, 581.24kb.
- Ударственными, научными и общественными организациями по анализу ситуации с загрязнением, 1224.32kb.
- Реферат на тему: пути загрязнения продовольственного сырья тяжелыми металлами, 407.26kb.
- «Химическое загрязнение почв» Общая трудоемкость дисциплины составляет, 25.84kb.
- Методика создания базы данных электронной модели рельефа. Современное состояние и экологические, 71.06kb.
- Глобальные последствия загрязнения окружающей среды, 130.02kb.
- Б. Б. Прохоров Экология человека: понятийно-терминологический, 2497.75kb.
- «Формы загрязнения природной среды. Загрязнители атмосферы, гидросферы, литосферы., 361.41kb.
- Экология и устойчивое развитие россии, 516.92kb.
- Экологические перспективы использования природных росторегуляторов для предотвращения, 394.04kb.
В подросте на территории исследуемого объекта произрастают ясень, клен, липа, которые в целом возобновляются успешно. Дуб в данных условиях семенным путём не возобновляется. Это, отчасти, можно объяснить тем, что под влиянием компонентов шлакоотвала, сформировалась почвенная среда, которая препятствует возобновлению дуба.
Отмечено отрицательное влияние шлакоотвала на естественное лесовозобновление: на удалении до 300 м оно отсутствует полностью; на расстоянии около 500 м происходит постепенное увеличение густоты подроста. Зависимость увеличения густоты подроста по мере удаления от шлакоотвала можно выразить уравнением прямой y = 6,7209x - 725,88 с коэффициентом аппроксимации R2 = 0,7357. Густота подлеска, наоборот, снижается по мере удаления от шлакоотвала, т.к. более здоровый древостой и подрост являются мощным конкурентом подлеска. Эту зависимость можно выразить уравнением параболы: y = -4E - 06x3 + 0,0196x2 - 32,53x + 17434 с коэффициентом аппроксимации R2 = 0,9564.
Сравнение полученных данных с результатами исследования 1999 г. показало, что на ПП, расположенных в зоне до 300 м от шлакоотвала, увеличилась густота подлеска, а на ПП далее 300 м улучшилось состояние подроста и увеличилась его густота, что объясняется ослаблением корневой конкуренции в связи повышенным отпадом деревьев основного полога.
Анализ естественного возобновления показывает сукцессию дубовых насаждений на липово-кленово-ясеневые, что с одной стороны, повышает биоразнообразие урочища и его устойчивость к воздействию шлакоотвала и других техногенных факторов, с другой – снижает его лесоводственно-экономическую значимость.
Содержание тяжёлых металлов в растениях и некоторые приемы
повышения экологической безопасности плодов и ягод
При попадании в почву металлы интенсивно поглощаются растениями, что показывают результаты анализа образцов древесины дуба черешчатого, ясеня обыкновенного и берёзы повислой, взятых на различном удалении (50 и 1000 м) в восточном направлении от шлакоотвала. Исследовались также листья дуба черешчатого, взятые с модельных деревьев, расположенных в 20 – 70 м на юго-восток от шлакоотвала (рисунок 11).
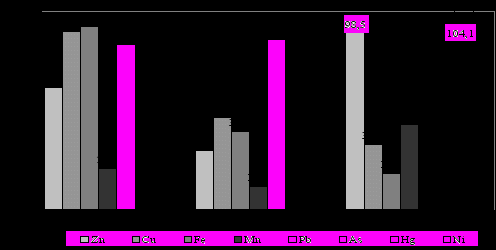
Рисунок 11 – Относительное изменение содержания металлов в древесине основных лесообразующих пород на расстоянии 1000 м от шлакоотвала
(за 100% взято содержание тяжёлых металлов на расстоянии 50 м)
Сравнительный анализ содержания металлов в древесине перечисленных видов показывает снижение их содержания на удалении 1000 м от шлакоотвала по сравнению с 50-метровой зоной в среднем, на 24,7 % для дуба, на 52,9 % для ясеня и на 56,6 % для берёзы.
Наличие разнообразных путей поступления тяжёлых металлов в растение предполагает существование двух ведущих факторов формирования элементного химического состава растений: генетического и экологического. Долевое участие каждого фактора меняется в зависимости от изменений условий среды. Наши исследования показали значительное влияние экологического и генетического факторов на содержание микроэлементов в плодах и листьях ягодных растений.
Листья смородины чёрной содержат в несколько раз больше Pb, Ni, Zn, Fe и Cu по сравнению с листьями малины и крыжовника (таблица 8). При этом содержание микроэлементов в листьях ягодных культур зависело от агрофона. Отмечено статистически достоверное влияние минеральных удобрений на накопление в листьях смородины чёрной Pb, Ni, Zn и Fe, и на накопление в листьях малины Ni и Zn.
Таблица 8 – Содержание микроэлементов в листьях ягодных культур (средние данные за 2005…2008 гг.).
| Культура | Вариант | Содержание микроэлементов в листьях, мг/кг сырой массы. | ||||
| Pb | Ni | Zn | Cu | Fe | ||
| Смородина чёрная | Контроль | 2,95 | 8,46 | 9,06 | 17,72 | 14,10 |
| N90Р90К90 | 3,26 | 6,09 | 10,85 | 16,70 | 39,84 | |
| Малина | Контроль | 0,68 | 1,55 | 5,96 | 3,21 | 3,36 |
| N90Р90К90 | 0,83 | 4,49 | 3,98 | 0,93 | 0,93 | |
| Крыжовник ЭЛС 24-15-21 | Контроль | 0,74 | 1,01 | 3,64 | 1,16 | 3,04 |
| N90Р90К90 | 0,71 | 1,08 | 4,22 | 1,00 | 3,06 | |
| Крыжовник ЭЛС 24-15-2 | Контроль | 0,9 | 0,83 | 3,42 | 1,2 | 1,02 |
| N90Р90К90 | 1,04 | 0,76 | 5,66 | 1,29 | 3,78 | |
Внесение цеолита в возрастающих дозах приводило к достоверно более низкому содержанию Pb и Ni в листьях смородины чёрной (рисунок 12), малины и крыжовника. Эти токсичные элементы поступают в растения преимущественно пассивным путём, и у всех изучаемых культур содержание Pb и Ni в листьях изменяется в соответствии с содержанием доступных форм в почве. Коэффициенты корреляции между содержанием Ni в листьях и подвижных форм Ni в почве были: для смородины – 0,7, для малины – 0,9, для крыжовника – 0,5. Корреляция между содержанием Pb в листьях и в почве: для малины r = 0,5, для крыжовника r = 0,82.
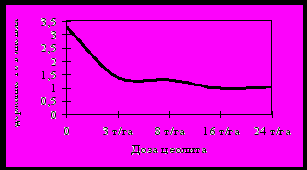
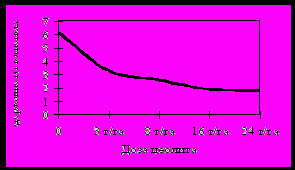
Рисунок 12 – Содержание Pb и Ni в листьях смородины чёрной
(мг/кг сырой массы) при внесении возрастающих доз цеолита.
Накопление биогенных элементов Zn и Cu в листьях ягодных культур при снижении содержания в почве доступных форм этих элементов при внесении возрастающих доз цеолита было специфичным для каждой культуры (рисунок 13). Увеличение содержания в листьях ягодных культур биогенных микроэлементов при снижении содержания в почве их доступных соединений может свидетельствовать о задействовании механизмов активного метаболического поглощения.
Таким образом, содержание токсичных микроэлементов в листьях ягодных культур определяется преимущественно экологическим фактором – содержанием в почве доступных форм этих элементов, а в поступлении в листья ягодных растений биогенных микроэлементов значимую роль играет генетический фактор.
Конечной целью исследований является оптимизация микроэлементного состава ягодной продукции. В таблице 9 приведены данные о содержании микроэлементов в плодах ягодных культур в условиях опыта, а также ПДК микроэлементов в плодах и ягодах и средние фоновые значения содержания микроэлементов в изучаемых культурах. Хотя содержание исследуемых металлов в плодах не превышает ПДК, присутствие Ni и Pb, изменение доли Zn, Cu и Fe по сравнению со средним фоновым уровнем свидетельствует о влиянии на минеральный состав внешних техногенных и агрогенных факторов и необходимости контроля за содержанием токсичных элементов.
Наибольшее содержание Pb отмечено в плодах крыжовника, Ni – в плодах малины. В то же время, малина характеризуется и наибольшим содержанием ценных биогенных микроэлементов – Fe и Zn.(Таблица 9).
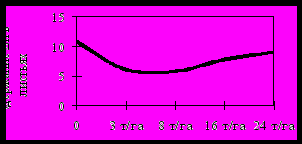
А Б
В Г
Д Е
А, Б – смородина чёрная; В, Г – малина;
Д

 , Е – крыжовник (форма ЭЛС 24-15-21 форма ЭЛС 24-15-2 )
, Е – крыжовник (форма ЭЛС 24-15-21 форма ЭЛС 24-15-2 )Рисунок 13 – Содержание Zn и Cu в листьях ягодных культур
при внесении возрастающих доз цеолита (мг/кг сырой массы);
Таблица 9 – Содержание микроэлементов в плодах ягодных культур
(средние данные за 2005…2008 гг.).
| Культура | Вариант | Содержание микроэлементов в плодах, мг/кг сырой массы. | ||||
| Pb | Ni | Zn | Cu | Fe | ||
| Смородина чёрная | Контроль | 0,025 | 0,15 | 0,48 | 0,26 | 0,32 |
| N90Р90К90 | 0,025 | 0,20 | 0,58 | 0,35 | 0,63 | |
| Средние фоновые | - | - | 0,8 | 0,8 | 8,4 | |
| Малина | Контроль | 0,016 | 0,354 | 1,845 | 0,259 | 3,77 |
| N90Р90К90 | 0,014 | 0,342 | 1,353 | 0,243 | 3,93 | |
| Средние фоновые | - | 0,06 | 2,0 | 1,7 | 12,0 | |
| Крыжовник ЭЛС 24-15-21 | Контроль | 0,053 | 0,101 | 0,845 | 0,160 | 0,489 |
| N90Р90К90 | 0,041 | 0,146 | 0,802 | 0,131 | 0,421 | |
| Крыжовник ЭЛС 24-15-2 | Контроль | 0,058 | 0,164 | 0,891 | 0,147 | 0,403 |
| N90Р90К90 | 0,045 | 0,206 | 0,856 | 0,123 | 0,450 | |
| | Средние фоновые | - | - | 0,9 | 1,3 | 8,0 |
| ПДК для плодов и ягод | 0,4 | 0,5 | 10,0 | 5,0 | 50,0 | |
Суммарный показатель накопления Pb, Ni, Zn и Cu составил 0,915 – для смородины чёрной, 2,47 – для малины, 1,16 и 1,26 – соответственно, для форм крыжовника ЭЛС 24-15-21 и ЭЛС 24-15-2. Таким образом, в условиях наших опытов малина имеет наибольший экологический риск накопления ТМ в продукции.
Действие физиологических барьеров, препятствующих поступлению ТМ в генеративные органы, у разных культур проявляется неодинаково. Так смородина, имевшая самое высокое содержание ТМ в листьях (Таблица 8), содержала ТМ в ягодах в 30…50 раз меньше. Содержание Pb в плодах малины было в 44 раза меньше, чем в листьях. Содержание Ni, Zn и Cu – соответственно в 4, 3 и 12 раз меньше. В то же время, плоды и листья малины содержали почти одинаковое количество железа. (Таблицы 8 и 9).
Закономерности поступления Pb и Ni в плоды ягодных культур сходны с поступлением этих элементов в листья. Отмечена достоверная корреляция между содержанием Pb и Ni в плодах и листьях малины (rPb = 0,83 и
rNi = 0,76), крыжовника (rPb = 0,96 и rNi = 0,94), смородины чёрной (rNi= 0,77). Содержание Pb и Ni в плодах при внесении цеолита снижалось на 20…70% в зависимости от культуры.
Изменение содержания Zn и Cu в плодах при внесении возрастающих доз цеолита было специфичным для каждой культуры (таблицы 10 и 11).
Закономерности поступления Zn и Cu в ягоды смородины чёрной сходны с поступлением Zn и Cu в листья, что подтверждается корреляцией между содержанием микроэлементов в листьях и ягодах: rCu =0,8; rZn =0,5. Содержание Zn и Cu в ягодах смородины чёрной было связано и с содержанием подвижных форм ТМ в почве. Достоверные коэффициенты корреляции между содержанием этих элементов в почве и в ягодах составили: rCu =0,78;
rZn = 0,73. Таким образом, накопление Zn и Cu в ягодах смородины чёрной происходит при участии как генетического, так и экологического факторов.
В плодах малины минимальное содержание Zn и Cu отмечено в варианте с внесением 10 т/га цеолита на фоне N90Р90К90. (Таблица 10).
Таблица 10 – Содержание Zn и Cu в плодах смородины чёрной и малины
(мг/кг сырой массы).
| Смородина чёрная | Малина | ||||
| Вариант | Zn | Cu | Вариант | Zn | Cu |
| Контроль | 0,48 | 0,26 | Контроль | 1,854 | 0,259 |
| N90Р90К90 | 0,58 | 0,35 | N90Р90К90 | 1,353 | 0,243 |
| N90Р90К90+ Цеолит 3 т/га | 0,15 | 0,18 | N90Р90К90+ Цеолит 2 т/га | 1,491 | 0,192 |
| N90Р90К90+ Цеолит 8 т/га | 0,23 | 0,20 | N90Р90К90+ Цеолит 10 т/га | 1,073 | 0,167 |
| N90Р90К90+ Цеолит 16 т/га | 0,34 | 0,25 | N90Р90К90+ Цеолит 20 т/га | 1,230 | 0,176 |
| N90Р90К90+ Цеолит 24 т/га | 0,39 | 0,40 | N90Р90К90+ Цеолит 30 т/га | 1,641 | 0,174 |
| НСР 0,05 | 0,09 | 0,10 | НСР 0,05 | 0,110 | 0,030 |
Плоды крыжовника в вариантах с внесением цеолита содержали Zn достоверно меньше, чем в контрольном варианте (таблица 11). Отмечена высокая положительная корреляция (r = 0,91…0,94) между содержанием подвижных форм Zn в слое почвы 20…40 см и содержанием этого элемента в плодах обеих изучаемых форм.
Таблица 11 – Содержание Zn и Cu в плодах крыжовника (мг/кг сырой массы).
| ЭЛС 24-15-21 | ЭЛС 24-15-2 | ||||
| Вариант | Zn | Cu | Вариант | Zn | Cu |
| Контроль | 0,845 | 0,160 | Контроль | 0,891 | 0,147 |
| N90Р90К90 | 0,802 | 0,131 | N90Р90К90 | 0,856 | 0,123 |
| N90Р90К90+ Цеолит 8 т/га | 0,211 | 0,081 | N90Р90К90+ Цеолит 8 т/га | 0,243 | 0,153 |
| N90Р90К90+ Цеолит 16 т/га | 0,187 | 0,259 | N90Р90К90+ Цеолит 16 т/га | 0,306 | 0,214 |
| N90Р90К90+ Цеолит 24 т/га | 0,048 | 0,128 | N90Р90К90+ Цеолит 24 т/га | 0,064 | 0,155 |
| НСР 0,05 | 0,130 | 0,080 | НСР 0,05 | 0,080 | 0,010 |
Внесение 16 т/га цеолита на фоне N90Р90К90 способствовало достоверно более высокому содержанию Cu в плодах крыжовника по сравнению с фоном. Имела место отрицательная корреляция (r = -0,9) между содержанием Cu в плодах крыжовника и содержанием подвижных форм Cu в слое почвы 20…40 см. Всё это свидетельствует о преимущественно генетическом контроле поступления Cu в плоды крыжовника в экологических условиях опытного участка.
Под влиянием цеолита изменилось соотношение микроэлементов в плодах – суммарная доля токсичных элементов свинца и никеля снизилась на 20…50% в зависимости от культуры. Суммарная доля биогенных элементов в ягодах смородины чёрной возросла в основном за счет увеличения содержания важного в биологическом и пищевом плане железа. В плодах крыжовника суммарная доля биогенных элементов возросла за счёт железа и меди.
Минимальные значения суммарного показателя накопления (СПН) Pb, Ni, Zn и Cu в плодах ягодных культур отмечены в следующих вариантах: в опыте со смородиной чёрной – при внесении 3 т/га цеолита на фоне N90Р90К90, в опыте с малиной – при внесении 10 т/га цеолита на фоне N90Р90К90, в опыте с крыжовником – при внесении 24 т/га цеолита на фоне N90Р90К90.
Анализ содержания фитогормонов в листьях смородины чёрной сорта Кипиана на территории ВНИИСПК показал, что внесение под эту культуру N90Р90К90 существенно (в 4 раза) увеличивает содержание ИУК на фоне повышения соотношения ИУК/АБК (6,80 против 1,86 на контроле). В варианте с совместным применением N90Р90К90 и цеолита, несмотря на максимальное содержание ИУК, соотношение ИУК/АБК было на уровне контроля (2,14 относительно 1,86 в контроле). В условиях техногенного загрязнения в варианте N90Р90К90 + цеолит отмечено меньшее снижение ИУК (в 1,36 раза), чем в варианте с N90Р90К90 без цеолита (в 3,5 раза) по сравнению с их бесстрессовыми вариантами. В варианте с цеолитом количество АБК в листьях снизилось на 27%. В результате соотношение ИУК/АБК в растениях, произрастающих на территории техногенного загрязнения, при совместном внесении под культуру N90Р90К90 + цеолит было на уровне такового соотношения фитогормонов в варианте N90Р90К90 + цеолит на территории ВНИИСПК. Аналогичные данные в условиях стресса получены и по гибберелловой кислоте (ГА3). Возделывание растений на фоне совместного внесения в почву N90Р90К90 + цеолит в условиях интоксикации ТМ не повлияло на уровень ГА3, тогда как в варианте с N90Р90К90 количество данного гормона снизилось на 26%.
Таким образом, проведённый анализ показал, что в условиях интоксикации ТМ в варианте N90Р90К90 + цеолит гомеостаз фитогормонов был на относительно постоянном уровне, что может служить указанием на протекторное влияние используемого мелиоранта.
Биологически активные вещества могут изменять реакцию растений на стрессоры. В связи с этим особый интерес вызывает циркон – природный регулятор эндогенного фенольного действия, на основе гидроксикоричных кислот. По рекомендациям производителей циркон считается корнеобразователем и индуктором цветения, способствует формированию устойчивости к ряду грибных заболеваний. В связи с этим представляло интерес изучить возможность протекторного воздействия циркона на процесс липопероксидации при интоксикации растений земляники (сорт Рубиновый кулон) ионами меди.
Проведённые анализы показали, что внесение в почву 2 ПДК меди в 2,7 раза увеличило содержание исследуемого металла в листьях земляники по отношению к контрольным растениям. На фоне интоксикации растений медью обработка цирконом несколько снизила накопление исследуемого металла. Содержание ионов меди в данном варианте только в 2 раза превышало контроль.
Значительное накопление меди в растениях в варианте с 2 ПДК на 32,2 % повысило активность пероксидазы в листьях. По-видимому, увеличение активности фермента связано с наличием высокого уровня гидроперекисей (содержание последних на 50,3 % выше контроля). При этом коэффициент корреляции между активностью фермента и содержанием гидроперекисей составил 0,92.
На фоне двукратного увеличения содержания меди в листьях земляники обработка растений цирконом в варианте 2 ПДК меди + циркон на 30,8 % вызвала повышение активности пероксидазы по отношению к контролю, что практически не составляло отличия от увеличения активности фермента в варианте с 2 ПДК токсиканта. Однако образование гидроперекисей в данном варианте было несколько ниже, чем в варианте с чистым токсикантом (38 % против 50,3 %, соответственно, относительно контроля). Более низкое содержание гидроперекисей в варианте 2 ПДК меди + циркон, возможно, связано с активацией гидроксикоричными кислотами другого антиоксидантного фермента – супероксиддисмутазы (СОД). Известно, что в результате взаимодействия СОД с О2- происходит образование перекисей (Mitteler, 2002). Дальнейшая реутилизация перекисей идет с участием каталазно-пероксидазной системы. Следует отметить, что обработка цирконом растений земляники, растущей на почве без внесения меди, достоверно не повлияла ни на активность фермента, ни на образование гидроперекисей.
Вызванные изменения в содержании катионов меди, гидроперекисей и активности антиоксидантного фермента в листьях растений, в конечном итоге сказались и на интенсивности перекисного окисления липидов мембран. Содержание малонового диальдегида при интоксикации земляники ионами меди было на 63,84 % выше контроля. Обработка растений цирконом не только несколько снижала накопление ионов меди и образование гидроперекисей в варианте 2 ПДК меди + циркон, но и препятствовала интенсификации ПОЛ. Так, количество конечного продукта липопероксидации – МДА в данном варианте было выше контроля только на 45,2 %.
Таким образом, используемый регулятор роста циркон сдерживал накопление ионов меди в листьях земляники при её токсичном содержании в почве и регулировал перекисный гомеостаз растений.