
Полиморфизм и функциональные свойства гена lmp1 вируса эпштейна-барр у больных лимфопролиферативными заболеваниями в россии 14. 00. 14. онкология
| Вид материала | Автореферат |
- «Всероссийский центр экстренной и радиационной медицины имени А. М. Никифорова», 382.53kb.
- Приказ 17 апреля 1998 г. N 125 о стандартах (протоколах) диагностики и лечения больных, 716.63kb.
- Операционно-анестезиологический риск и пути его снижения у гериатрического контингента, 934.6kb.
- Программа «Антисмог» в санатории «Валуево», 20.86kb.
- Николаева Мария Андреевна, доктор технических наук, профессор, заведующий кафедрой, 367.89kb.
- Календарно-тематический план лекций по ортопедической стоматологии для студентов Укурса, 23.79kb.
- Генетический полиморфизм cd14, il4, tnf и fcgr2A у больных гриппом a h1N1 в Забайкальском, 62.14kb.
- Задачи: Оценить влияние комплекса артролон на динамику клинической картины у пациентов, 672.18kb.
- Задачи : Оценить влияние комплекса кардиомакс на динамику клинической картины у больных, 60.17kb.
- Функциональные свойства паремий-трансформов в англоязычном публицистическом дискурсе, 736.64kb.
19

На правах рукописи
СМИРНОВА КСЕНИЯ ВАЛЕРЬЕВНА
ПОЛИМОРФИЗМ И ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА ГЕНА LMP1 ВИРУСА ЭПШТЕЙНА-БАРР У БОЛЬНЫХ ЛИМФОПРОЛИФЕРАТИВНЫМИ ЗАБОЛЕВАНИЯМИ В РОССИИ
14.00.14. – онкология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
МОСКВА - 2009

Работа выполнена в лаборатории вирусного канцерогенеза НИИ канцерогенеза
Учреждения Российской академии медицинских наук
Российского онкологического научного центра им. Н.Н. Блохина РАМН
(директор – академик РАН и РАМН, профессор М.И. Давыдов)
Научный руководитель:
доктор медицинских наук, профессор В.Э. Гурцевич
Официальные оппоненты:
доктор биологических наук, профессор,
член-корр. РАМН Ф.Л. Киселев
доктор биологических наук Л.Э.Завалишина
Ведущая организация:
ГНЦ Институт иммунологии ФМБА РФ, г. Москва
Защита состоится « ____ » ________________ 2009 г. в _______ часов на заседании диссертационного ученого совета (Д.001.017.01) Российского онкологического научного центра им. Н.Н. Блохина РАМН по адресу 115478, г. Москва, Каширское шоссе, 24.
С диссертацией можно ознакомиться в библиотеке
РОНЦ им. Н.Н. Блохина РАМН
Автореферат разослан « ____ » _______________ 2009 г.
Ученый секретарь диссертационного совета,
доктор медицинских наук, профессор Ю.В. Шишкин
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Латентный мембранный белок 1 (LMP1), являясь онкогеном вируса Эпштейна-Барр (ВЭБ), представляет собой интегральный мембранный белок, кодируемый одноименным геном LMP1, экспрессия которого приводит к трансформации клеток крысиных фибробластов и B-лимофцитов in vitro [Kaye et al., 1993]. LMP1 играет также ключевую роль в развитии лимфом у трансгенных мышей [Kulwichit et al., 1998]. В состав белка входят два терминальных цитоплазматических и шесть трансмембранных доменов [Fennewald et al., 1984]. Длинный C-терминальный цитоплазматический домен ответственен за индукцию множества клеточных сигнальных каскадов, приводящих к активации NF-kB, c-Jun N-терминальной киназы (JNK), STAT 1/3, p38 MAPK [Wu et al., 2006, Eliopoulos et al., 1998, 1999, Dawson et al., 2003, Lam et al., 2003]. Основной функцией короткого N-терминального, а также трансмембранных доменов, является интеграция молекул LMP1 в клеточную мембрану и их агрегация между собой [Coffin et al., 2001].
Филогенетический анализ вариантов LMP1, выделенных из опухолевой ткани больных ВЭБ-ассоциированными патологиями в различных географических регионах, выявил аккумуляцию ряда мутаций в этом гене, что свидетельствует о высокой степени его полиморфизма. В настоящее время рассматривается три основных механизма изменчивости LMP1 – это возникновение точечных мутаций, ведущих к замене отдельных аминокислот; образование делеций и дупликаций; и гомологичная рекомбинация как следствие ко-инфекции лимфоидных или эпителиальных клеток двумя различными штаммами ВЭБ [Walling et al., 1999]. Исследование отдельных мутаций этого гена выявило способность некоторых из них влиять на биологические свойства молекулы LMP1, что, вероятно, играет важную роль в этиопатогенезе заболеваний, вызываемых ВЭБ. Наибольший интерес представляют мутации C-терминального цитоплазматического домена, которые, как показано ранее, влияют на иммуногенность и время полужизни LMP1, что в конечном итоге способствует усилению его трансформирующего воздействия на клетку [Hu et al., 1991, Edwards et al., 1999, Павлиш и др., 2008]. Не менее важную роль в изменениях свойств белка играют и точечные мутации, локализованные в трансмембранном домене LMP1, при этом показано, что некоторые из них приводят к значительному снижению цитотоксического действия вирусного белка на клетку, что, возможно, влияет на трансформирующий потенциал этого онкобелка [Nitta et al., 2004].
Генетические перестройки гена LMP1 характеризуют изоляты ВЭБ, полученные от больных различными лимфопролиферативными заболеваниями в различных странах мира. Поиск и молекулярно-генетический анализ таких изолятов в неэндемичной зоне, а именно на территории России, представляется а к т у а л ь н о й научной и практической задачей, которая позволит получить новые экспериментальные данные о трансформирующих свойствах гена LMP1 и выяснить роль новых изолятов ВЭБ в возникновении различных лимфопролиферативных заболеваний, как доброкачественных, так и злокачественных.
Цель и задачи исследования
Цель настоящей диссертационной работы заключалась в изучении основных функциональных характеристик доминирующих вариантов LMP1 ВЭБ, персистирующих среди больных лимфопролиферативными заболеваниями в России.
Для решения поставленных целей определены следующие задачи:
1. Провести секвенирование и филогенетический анализ вариантов LMP1, амплифицированных из клинических образцов, полученных от больных различными лимфопролиферативными заболеваниями;
2. Клонировать наиболее распространенные среди населения России минимально дивергентные варианты LMP1 в ретровирусный вектор;
3. Получить клеточные линии, стабильно экспрессирующие варианты LMP1, на основе клеточной линии Rat-1 (крысиных фибробластов), трансфицированной LMP1-реторовирусными плазмидами;
4. Изучить морфологические изменения полученных клеточных линий, экспрессирующих варианты LMP1 российского происхождения;
5. Оценить активацию/супрессию основных сигнальных путей (NF-kB и АР-1), а также выявить влияние на накопление активных форм азота, в клеточных линиях, экспрессирующих низкодивергентные варианты LMP1.
Научная новизна и практическая значимость исследования
В представленной работе с помощью секвенирования и филогенетического анализа вариантов LMP1, изолированных из тканей российских больных ВЭБ-ассоциированными лимфопролиферативными заболеваниями, в частности инфекционным мононуклеозом (ИМ), лимфомой Ходжкина (ЛХ) и неходжкинскими лимфомами (НХЛ), доказано преобладание монофилетичеcкой группы, относящейся к низкодивергентному штамму LMP1-В95.8b. Учитывая происхождение изучаемых вариантов LMP1 из тканей больных доброкачественными (ИМ) и злокачественными (ЛХ и НХЛ) заболеваниями, представляло интерес сравнить их функциональные свойства с таковыми для высокодивергентного варианта LMP1-Cao, обладающего выраженным трансформирующим потенциалом, а также прототипного варианта LMP1-B95.8 с невысокой трансформирующей активностью. С этой целью нами впервые проведен анализ активации ключевых транскрипционных факторов NF-κB и AP-1, а также индукции iNOS и уровня внутриклеточного накопления NO, вариантами LMP1 с минимальным набором мутаций с одной стороны, и указанными контрольными вариантами белка с другой.
П р а к т и ч е с к о е з н а ч е н и е заключается в том, что проведенный анализ нуклеотидных последовательностей вариантов полноразмерного гена LMP1 ВЭБ выявил конкретные генетические отличия между изолятами LMP1, амплифицированными из тканей больных различными злокачественными и доброкачественными лимфопролиферативными заболеваниями. Обнаруженные функциональные отличия низкодивергентных вариантов LMP1 помогут расширить понимание сложного комплекса молекулярно-биологических, вирусологических и иммунологических закономерностей, которые лежат в основе патогенеза ВЭБ-ассоциированных заболеваний, а также в разработке новых подходов для ранней диагностики неоплазий человека.
Апробация работы
Диссертация апробирована 18 июня 2009 года на совместной научной конференции лабораторий вирусного канцерогенеза, иммунологии онкогенных вирусов, молекулярной биологии вирусов, регуляции клеточных и вирусных онкогенов, биохимии опухолей, методов скрининга канцерогенов НИИ Канцерогенеза РОНЦ им. Н.Н. Блохина РАМН. Материалы работы докладывались на 16-ой и 17-ой международных конференциях “AIDS, Cancer and Public Health” 2007, 2008 (Санкт-Петербург, Россия), Всероссийской научно-практической конференции “Отечественные противоопухолевые препараты” 2007, 2008, 2009 (Москва, Россия), 8-ой международной школе-конференции “Immunology and viral infection” 2007 (Москва, Россия), 5-ой Российско-немецкой конференции “Human herpesvirus infections” 2008 (Москва, Россия), третьем европейском конгрессе по вирусологии 2007 (Нюрнберг, Германия), 20-ом симпозиуме Европейской ассоциации по изучению рака (EACR) 2008 (Лион, Франция), пятой конференции «Фундаментальная онкология – Петровские чтения» 2009 (Санкт-Петербург, Россия).
Публикации
По материалам диссертации опубликовано 4 научные статьи и 13 тезисов докладов.
Структура и объем диссертации
Диссертация изложена на 117 страницах машинописного текста, содержит 17 рисунков и 4 таблицы. Состоит из глав: Введение, Обзор литературы, Материалы и методы, Результаты исследования, Обсуждение, Выводы, Список литературы. Список используемой литературы содержит 248 литературных источников, из них 7 отечественных и 241 зарубежных.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы
Клинический материал. Материалом для исследования служили образцы крови, биопсийный материал опухолей и смывы из ротоглотки от 13 больных лимфомой Ходжкина (ЛХ), 13 больных неходжкинскими лимфомами (НХЛ), проходивших лечение в Российском Онкологическом Научном Центре им. Н.Н.Блохина РАМН (РОНЦ РАМН), а также 10 образцов крови больных инфекционным мононуклеозом (ИМ) из детской клинической больницы №38 г. Москвы.
Плазмиды. При проведении исследования использовали векторные конструкции pSG5-LMP1-B95-8 и pSG5-LMP1-Cao на основе эукариотического экспрессирующего вектора pSG5, содержащего ранний промотор и энхансер SV40, а также β-глобиновый интрон, обеспечивающий процесс сплайсинга клонированных эукариотических генов. Исследуемые варианты гена LMP1 с использованием соответствующих эндонуклеаз рестрикции переклонировали из системы pGEM-T Easy («Promega», США) в эукариотический экспрессирующий вектор pSG5 и в ретровирусный вектор pBabe-puro (pBabe). Для анализа активации транскрипционного фактора NF-kB использовали репортерную плазмиду kB-ConA-Luc, предоставленную F. Grasser (Хомбург, Германия). Для проведения анализа активации JNK сигнального пути использовали jun2-люциферазную репортерную плазмиду (АР1-Luc), любезно предоставленную M. Rowe (Кардиф, Англия).
Клеточные линии. В работе использовали клеточную линию Phi-NX-Ampho (Phoenix-A), дериват эмбриональных клеток почки человека HEK293, а также крысиные фибробласты Rat-1.
Моноклональные антитела. В ходе проведения исследования применяли моноклональные антитела к белку LMP1 – S12, индуцибельной форме NO-синтазы (NOS-20), а также специфические антитела к β-катенину.
Методы. При проведении исследований использовали следующие методы: выделение ДНК, ПЦР-анализ, филогенетический анализ, электрофорез нуклеиновых кислот, трансформация бактериальных штаммов, клонирование ДНК, трансфекция, трансдукция, аналитический электрофорез белков, иммуноблотинг, денситометрический анализ, люциферазный анализ, нитрат/нитритный анализ, статистическая обработка полученных данных осуществлялась с помощью программы GraphPad Prism ver. 5.0.
Результаты исследования
1. Филогенетический анализ вариантов LMP1, изолированных от российских больных ВЭБ-ассоциированными заболеваниями
Используя клинический материал российских больных ЛХ, НХЛ и ИМ, нами проведена амплификация и секвенирование полноразмерных вариантов гена LMP1 (табл. 1). Дальнейший филогенетический анализ показал, что исследуемые варианты вирусного онкобелка представляют собой гетерогенную популяцию, в состав которой входят варианты с варьирующим числом мутаций в опухолевой и нормальной тканях вне зависимости от имеющегося у больного типа патологии. В этих вариантах LMP1 обнаружены мутации, характерные для таких высокодивергентных штаммов, выделяемых в классификации Edwards с соав. (1999) [Edwards et al., 1999], как Ch1 (China 1), Ch2 (China 2), NC (New York City), Med+ (Mediterranean +), Med- (Mediterranean -). Абсолютно доминирующей, однако, была монофилетическая группа, отмеченная в классификации Sandvej с соавт., как штамм LMP1-В95.8b [Sandvej et al., 1997] (рис. 1). Для представителей данной группы характерен низкий уровень дивергенции (3-4 критические замены а.к.). Среди точечных замен аминокислот абсолютно превалирующими в этих вариантах являются замены изолейцина на лейцин в 85 положении (I85L), фенилаланина на тирозин в 106 положении (F106Y), глутаминовой кислоты на глутамин в 328 положении (E328Q) и серина на треонин в 366 положении (S366T).
Для представителей группы LMP1-В95.8b также характерно отсутствие Сао-подобной делеции 30 п.н. и наличие «молчащей» замены Т на С в 356 кодоне LMP1. Не принадлежащие к этому штамму высокодивергентные варианты LMP1 российского происхождения (Ch1, Ch2, Med+, Med-, NC), выделяемые в классификации Edwards с соав., содержали альтернативную «молчащую» замену в 342 кодоне, где А замещен на Т [Edwards et al., 1999]. Интересно отметить, что ни один из исследованных нами вариантов не содержал обе указанные «молчащие» замены нуклеотидов, что, возможно, свидетельствует о существовании двух cамостоятельных дивергентных линий гена LMP1.
По-видимому, генетическая линия изолятов вируса, к которой относится штамм В95.8b, не претерпела серьёзных мутационных перестроек в отличие от более дивергентных и гетерогенных представителей другой линии, к которой принадлежат остальные варианты LMP1. Функциональное значение некоторых встречающихся мутаций (I85L, F106Y, E328Q, S366T), как показано ранее, проявляется в снижении цитотоксичности и усилении трансформирующей активности белка LMP1 [Nitta et al., 2004, Tang et al., 2003].
Таблица 1. Варианты LMP1 российского происхождения,
использованные для филогенетического анализа
| Лимфома Ходжкина | Неходжкинские лимфомы | Инфекционный мононуклеоз |
| ЛХ-95.1 | НХЛ-27.3 | ИМ-2685 |
| ЛХ-95.2 | НХЛ-27.2 | ИМ-2783 |
| ЛХ-38.1 | НХЛ-101.1 | ИМ-2746 |
| ЛХ-82.1 | НХЛ-104.1 | ИМ-2717 |
| ЛХ-73.1 | НХЛ-27.1 | ИМ-2718 |
| ЛХ-77.1 | НХЛ-100.1 | ИМ-2799 |
| ЛХ-36.3 | НХЛ-39.3 | ИМ-2620 |
| ЛХ-34.1 | НХЛ-28.3 | ИМ-2624 |
| ЛХ-102.1 | НХЛ-28.2 | ИМ-2623 |
| ЛХ-108.1 | НХЛ-28.1 | ИМ-2780 |
| ЛХ-46.1 | НХЛ-41.3 | |
| ЛХ-62.1 | НХЛ-91.1 | |
| ЛХ-32.1 | НХЛ-99.1 | |
| | | |
| Σ: 13; В95.8b – 7 (53,9%) | Σ: 13; В95.8b – 6 (46,2%) | Σ: 10; В95.8b – 5 (50,0%) |
| | | |
| Всего: 36 вариантов из них В95.8b – 18 (51,4%) | ||
    0.1 | LMP1-В95.8b |
Рис. 1. Филогенетическое древо, построенное методом «ближайших соседей» (neighbor-joining) по полноразмерным аминокислотным последовательностям вариантов LMP1 российского происхождения.
Другие точечные мутации, обнаруженные нами в исследуемых вариантах (D210E, G352S, W39C, L93V, A96T, I122L, S239M), ранее не описаны, однако некоторые из них локализуются вблизи функциональных доменов LMP1 и, возможно, могут оказывать влияние на свойства этой молекулы.
Учитывая доминирующее распространение в России изолятов ВЭБ с низкодивергентными вариантами LMP1, в том числе у больных злокачественными (ЛХ и НХЛ) и доброкачественными (ИМ) лимфопролиферативными заболеваниями, представлялось важным провести сравнительное изучение биологических свойств LMP1 российского происхождения, принадлежащих к группе В95.8b. С этой целью варианты LMP1 из опухолевой ткани больных ЛХ (ЛХ-34.1, ЛХ-46.1) и НХЛ (НХЛ-28.1, НХЛ-28.2 и НХЛ-41.3), а также периферической крови больных ИМ (ИМ-2623, ИМ-2624) были изучены на их способность образовывать колонии, а также активировать ключевые транскрипционные факторы NF-κB и АP-1, индуцибельную форму NO-синтазы (iNOS) и внутриклеточное накопление окиси азота (NO). Характеристика изучаемых вариантов LMP1 представлена в табл. 2.
Из этой таблицы видно, что варианты LMP1, а



 мплифицированные из опухолевой ткани, крови или смыва ротоглотки характеризовались незначительным набором мутаций, специфичных для группы В95.8b. Опухолевые клетки больных ЛХ и НХЛ экспрессировали белок LMP1, выявленный методом иммуногистохимии. Хотя титры антител к ВКА ВЭБ в крови этих больных были невысокими, в отличие от высоких титров вирус-специфических антител у больных ИМ.
мплифицированные из опухолевой ткани, крови или смыва ротоглотки характеризовались незначительным набором мутаций, специфичных для группы В95.8b. Опухолевые клетки больных ЛХ и НХЛ экспрессировали белок LMP1, выявленный методом иммуногистохимии. Хотя титры антител к ВКА ВЭБ в крови этих больных были невысокими, в отличие от высоких титров вирус-специфических антител у больных ИМ. Присутствие белкового продукта LMP1 в опухолевой ткани больных подтверждалось с помощью иммуногистохимического исследования (авидин-биотин-пероксидазным (АБП) методом) со специфическими для LMP1 моноклональными антителами S12 (рис. 2).
Таблица 2. Характеристика низкодивергентных вариантов LMP1, использованных для исследования их биологических свойств.
| Варианты LMP1 | Материал | Аминокислотные замены | Титры IgG антител к ВКА* |
| ЛХ-34.1 | опухоль | F106Y, D210E, E328Q, S366T | 1:160 |
| ЛХ-46.1 | опухоль | I85L, F106Y, E328Q, G352S, S366T | н.о. |
| НХЛ-28.1 НХЛ-28.2 | опухоль кровь | W39C, L93V, F106Y, E328Q, S366T F106Y, E328Q, S366T | 1:80 |
| НХЛ-41.3 | смыв рото- глотки | I85L, A96T, F106Y, E328Q, S366T | 1:40 |
| ИМ-2623 | кровь | I85L, F106Y, I122L, L306M, E328Q, S366T | 1:320 |
| ИМ-2624 | кровь | I85L, F106Y, S239M, E328Q, S366T | 1:1280 |
* - в реакции непрямой иммунофлуоресценции.
Сокращения: ЛХ – лимфома Ходжкина, НХЛ – неходжкинская лимфома, ИМ – инфекционный мононуклеоз, 1 - опухоль, 2 - кровь, 3 - смыв ротоглотки, ВКА – вирускапсидный антиген, н.о. - не определялось.
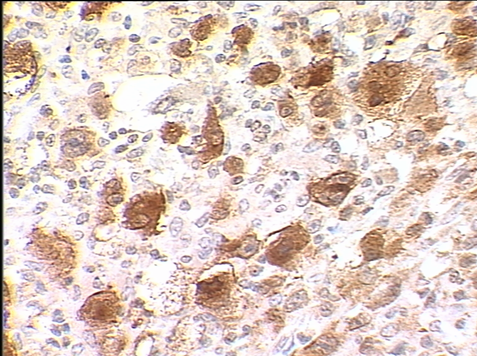
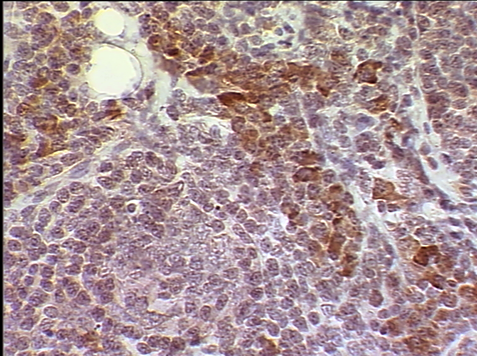

А
Б
Рис. 2. Иммуногистохимическое исследование на экспрессию белка LMP1. А – лимфома Ходжкина, нодулярный склероз. Опухолевые клетки с морфологией лакунарных клеток экспрессируют LMP1. Цитоплазматическая реакция. АБП-метод. х400. Б – лимфоплазмоцитарная лимфома (НХЛ). Опухолевые лимфоидные, лимфоплазмоцитарные и плазматические клетки, экспрессируют LMP1. Цитоплазматическая реакция. АБП-метод. х200.
2. Получение полноразмерных низкодивергентных вариантов LMP1 и клеточных линий, постоянно экспрессирующих исследуемые белки
2.1. Реконструкция полноразмерных вариантов LMP1
Наша стратегия получения выбранных полноразмерных вариантов гена LMP1 была основана на амплификации двух фрагментов каждого варианта гена – N-концевого (836 пн) и С-концевого (620 пн). При этом 5'-концевая область фрагмента 620 пн и 3'-концевая область фрагмента 836 пн частично перекрывались и имели общий внутренний сайт рестрикции для эндонуклеазы рестрикции MscI (BalI), что позволило в дальнейшем использовать его для реконструкции полноразмерных вариантов гена LMP1. Все манипуляции по объединению фрагментов в полноразмерный ген проводились в плазмидном векторе pGEM-T Easy («Promega»). Скрининг позитивных клонов вели по появлению «белых» колоний в присутствии стандартных концентраций ИПТГ и X-gal. Следует отметить, что клонирование было более эффективным при использовании XLblue-1 штамма E. coli, тогда как при работе со штаммом DH5α, наблюдали появление большого количества пустых «белых» колоний без вставки ПЦР-фрагмента.
Для изучения биологических свойств выбранных низкодивергентных LMP1 (в частности, для анализа сигнальной активности с использованием люциферазных репортерных векторов), исследуемые варианты гена клонировали в эукариотический экспрессирующий вектор pSG5. После необходимой рестрикционной проверки клонов, содержащих «вставку», выделяли препаративные количества плазмидных ДНК и использовали в дальнейших экспериментах.
2.2. Получение клеточных линий, стабильно экспрессирующих низкодивергентные варианты белка LMP1
С целью получения клеточных линий, стабильно экспрессирующих низкодивергентные варианты LMP1, нами проведено клонирование вариантов полноразмерных генов в ретровирусный вектор pBabe-puro по соответствующим сайтам. Для определения ориентации всех переклонированных вариантов гена LMP1 очищенные плазмидные ДНК рестрицировали по HindIII и Msp20I сайтам. При правильной ориентации вставок в векторе pBabe-puro вырезались фрагменты 0,5 kb (при клонировании по EcoRI сайту) и 2,5 kb (при клонировании по BamHI сайту) (рис.3).
Переклонирование полноразмерных вариантов гена из эукариотического экспрессирующего вектора в ретровирусный также проводили с целью исключения хорошо известного эффекта гиперэкспрессии LMP1, который наблюдается при трансфекции этого гена в составе эукариотических векторов в различные типы клеток и часто приводит к ошибочным результатам.
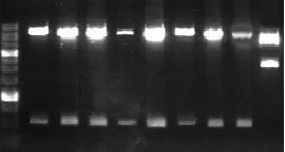
M
1
2
3
4
5
6
7
8
9
3kb
1kb

Рис. 3. Тест на ориентацию LMP1-содержащих фрагментов в pBabe-puro: 1 – LMP1-B95-8; 2 – ЛХ-34.1; 3 – ЛХ-46.1; 4 – НХЛ-28.1; 5 – НХЛ-28.2, 6 – НХЛ-41.3; 7 – ИМ-2623; 8 – ИМ-2624; 9 – LMP-Cao; М – маркер MW. С 1 по 8 – правильная ориентация EcoRI фрагментов; 9 – правильная ориентация BamHI фрагмента.
После наработки переклонированных в pBabe-puro низкодивергентных вариантов гена LMP1, а также его контрольных вариантов LMP1-B95-8 (низкотуморогенного) и LMP1-Cao (высокотуморогенного), проводилась их трансфекция в клеточную линию Phi-NX-Ampho (Phoenix-A, дериват клеток HEK293). Клетки данной линии содержат генетические конструкции (RSV-Gag-Pol-PolyA и CMV-env-PolyA), что позволяет обеспечить используемые в эксперименте векторные конструкты соответствующей «упаковкой», содержащей компоненты оболочки амфотропного вируса мышей. При транзиторной трансфекции клеток Phoenix-A ретровирусными векторами, несущими соответствующие вставки гена LMP1, формируется высокий титр вирусов (до 107 на мл), которые продуцируются трансфецированными клетками в культуральную среду. Далее полученными ретровирусными стоками мы трансдуцировали индикаторные клетки эмбриональных крысиных фибробластов Rat-1. Выбор этой клеточной линии, в первую очередь, был обусловлен чувствительностью этих клеток к трансформирующим свойствам LMP1 и их реактивностью к трансформирующему действию карбоксильного домена белка [Moorthy et al., 1993], а также результатами предыдущих исследований, выявивших их меньшую чувствительность к цитотоксическому действию молекулы LMP1 [Дидук и др., 2008]. Последующую селекцию клеток, несущих необходимые векторные конструкции, проводили в течение двух недель в присутствии пуромицина.
Экспрессию исследуемых вариантов LMP1 и их контрольных вариантов в трансдуцированных клетках Rat-1 проверяли с помощью иммуноблотинга со специфическими антителами к области повторов молекулы. Во всех трансдуцированных клеточных линиях нами обнаружены продукты экспрессии низкодивергентных вариантов LMP1, соответствующего размера (63 кДа). Как видно на рисунке 4, экспрессия белка во всех линиях происходит примерно на одинаковом уровне.
1
8
9
10

2
3
4
6
7
5
63 кДа

Рис. 4. Экспрессия LMP1 после трансдукции клеток Rat-1 (иммуноблотинг с мАТ S12): 1 – pBabe, 2 – LMP1-B95-8; 3 – ЛХ-34.1; 4 – ЛХ-46.1; 5 – НХЛ-28.1; 6 – НХЛ-28.2; 7 – НХЛ-41.3; 8 – ИМ-2623; 9 – ИМ-2624; 10 – LMP-Cao.
3. Морфологические изменения клеточных линий, экспрессирующих низкодивергентные варианты LMP1
Поскольку LMP1 является белком-онкогеном ВЭБ и блокирует контактное торможение фибробластов Rat-1, мы решили проверить влияние минимального набора мутаций в исследуемых вариантах белка на этот процесс. С помощью метода фазового контраста был проведен анализ способности к формированию фокусов трансформации клеточными линиями, постоянно экспрессирующими исследуемые варианты LMP1. Как видно из табл. 3, клетки, содержащие контрольный пустой вектор pBabe, при длительном культивировании не
Таблица 3. Количество фокусов трансформации, образуемое клеточными линиями, экспрессирующими низкодивергентные и контрольные варианты LMP1.
| Клеточные линии | Количество фокусов трансформации |
| Rat-1-pBabe-puro | 0 |
| Rat-1-pBabe-B95-8 | > 90 |
| Rat-1-pBabe-ЛХ-34.1 | >110 |
| Rat-1-pBabe-ЛХ-46.1 | > 100 |
| Rat-1-pBabe-НХЛ-28.1 | >110 |
| Rat-1-pBabe-НХЛ-28.2 | > 120 |
| Rat-1-pBabe-НХЛ-41.3 | > 100 |
| Rat-1-pBabe-ИМ-2623 | > 120 |
| Rat-1-pBabe-ИМ-2624 | > 100 |
| Rat-1-pBabe-Cao | > 250 |
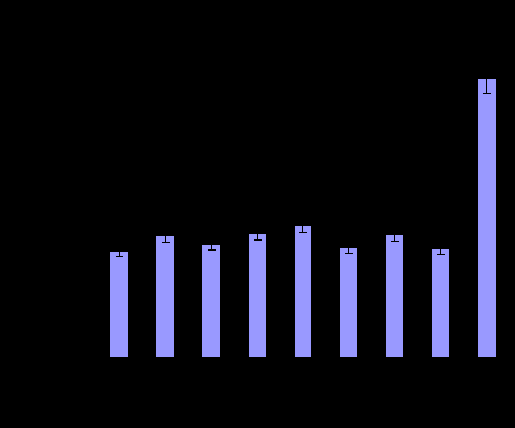
Рис. 5. Количество фокусов трансформации, образуемое клеточными линиями, экспрессирующими различные варианты LMP1.
образовывали фокусы трансформации, в то время как в остальных линиях при экспрессии вариантов LMP1 B-95.8, ЛХ-34.1, ЛХ-46.1, НЛХ-28.1, НЛХ-28.2, НЛХ-41.3, ИМ-2623 и ИМ-2624 формировались очаги многослойного роста разного размера. Клетки, экспрессирующие вариант LMP1-Сао, образовывали достоверно большее количество фокусов трансформации по сравнению с остальными линиями (рис. 5).
Для дальнейшего определения трансформирующих способностей клеточных линий, постоянно экспрессирующих низкодивергентные LMP1, а также контрольные варианты, мы анализировали их возможность к росту без подложки в жидком агаре. Эксперимент показал, что все исследуемые варианты при культивировании в жидком агаре образовывали колонии различного размера, при этом, так же как и в случае описанных выше экспериментов, клетки, экспрессирующие высокотуморогенный вариант LMP1-Cao образовывали намного большее количество колоний (табл. 4, рис. 6).
Исходя из вышесказанного, можно сделать вывод о влиянии даже ограниченного числа мутаций в молекуле LMP1, как в транмембранных, так и в цитоплазматическом доменах, на туморогенные свойства вариантов LMP1. Хотя необходимо констатировать, что этих мутаций все же недостаточно для проявления более агрессивных трансформирующих свойств, которые наблюдаются при экспрессии высокотуморогенного варианта LMP1-Cao.
4. Функциональный анализ низкодивергентных вариантов LMP1, полученных от больных лимфопролиферативными заболеваниями
4.1. Оценка функциональной способности вариантов LMP1 активировать транскрипционные факторы NF-κB и jun/AP-1
Как известно, экспрессия белка LMP1 приводит к активации множества клеточных генов, вовлеченных в процессы пролиферации, иммортализации и выживаемости клеток. Индукция многих из этих генов тесно связана с активацией таких ключевых транскрипционных факторов клетки, как NF-κB и AP-1.
Таблица 4. Количество колоний, образуемое клеточными линиями, экспрессирующими низкодивергентные и контрольные варианты LMP1 при росте без подложки.
| Клеточные линии | Количество колоний | |
| Rat-1-pBabe-puro | 0 | |
| Rat-1-pBabe-B95-8 | > 30 | |
| Rat-1-pBabe-ЛХ-34.1 | >40 | |
| Rat-1-pBabe-ЛХ-46.1 | > 35 | |
| Rat-1-pBabe-НХЛ-28.1 | >30 | |
| Rat-1-pBabe-НХЛ-28.2 | > 40 | |
| Rat-1-pBabe-НХЛ-41.3 | > 28 | |
| Rat-1-pBabe-ИМ-2623 | > 37 | |
| Rat-1-pBabe-ИМ-2624 | > 30 | |
| Rat-1-pBabe-Cao | > 50 | |
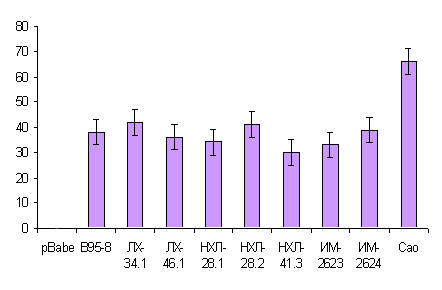
Рис. 6. Количество колоний, образуемое клеточными линиями, экспрессирующими различные варианты LMP1 при росте без подложки.
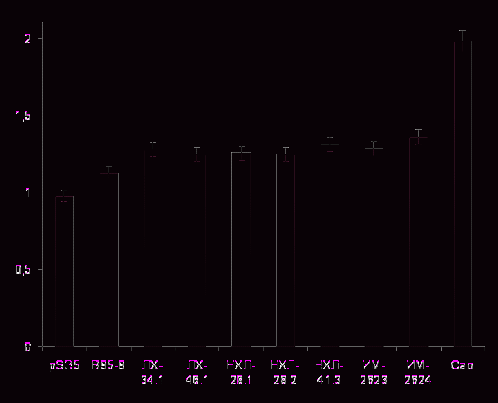
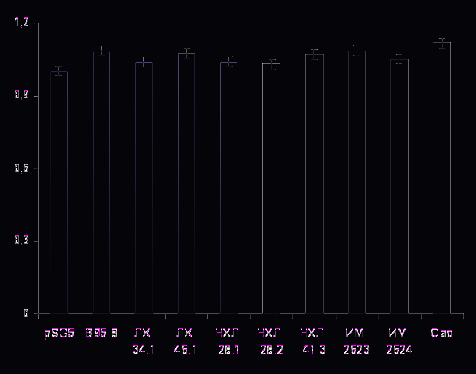
Используя соответствующие люциферазные репортерные векторы, мы проанализировали влияние низкодивергентных вариантов LMP1, полученных от российских больных ЛХ, НХЛ и ИМ, и двух контрольных вариантов LMP1-B95.8 и LMP1-Cao (прототипного и высокотуморогенного, соответственно) на уровни активации NF-κB и АP-1. Как следует из рисунка 7а, экспрессия исследуемых вариантов белка в клетках HEK293 приводит к активации NF-κB, уровень которой, однако, на 5-8% (p ≥ 0,05) выше такового, вызванного экспрессией прототипного варианта LMP1-B95.8. При этом значимых различий в активации этого транскрипционного фактора между самими изучаемыми низкодивергентными вариантами LMP1 выявить не удалось. В то же время, уровень активации NF-κB высокотуморогенным LMP1-Cao оказался в 1,83 раза выше таковых, вызванных прототипным вариантом LMP1-B95.8 и исследуемыми вариантами белка.
При изучении транскрипционного фактора jun/AP-1 также обнаружена способность у изучаемых вариантов LMP1 вызывать его активацию (рис. 7б). При этом экспрессия низкодивергентных LMP1, изолированных из опухолевой ткани больных ЛХ, НХЛ и периферической крови больных ИМ, а также экспрессия контрольных вариантов прототипного LMP1-B95.8 и высокотуморогенного LMP1-Сао, вызывала сходный уровень активации AP-1, что, вероятно, не отражает онкогенный потенциал исследуемых вариантов LMP1. При определении люциферазной активности наличие экспрессии LMP1 в анализируемых клеточных линиях подтверждали с помощью иммуноблотинга с использованием антител S12, специфичных к области повторов белка (рис.7в). Отсутствие значимых различий в активации AP-1 между изученными вариантами LMP1 различной степени дивергентности, отличающихся по набору мутаций, по-видимому, свидетельствует об отсутствии их критической роли в этом процессе. В то же время, существование природных вариантов LMP1, способных активировать этот транскрипционный фактор в независимости от замен в доменах этого белка наводит на предположение о существовании дополнительных факторов, влияющих на активацию АР-1.
Таким образом, на основании полученных данных можно сделать вывод о том, что минимальный набор мутаций в исследуемых вариантах вирусного белка, по-
видимому, не играет критической роли в изменении активности транскрипционных факторов NF-κB и AP-1.
А
Б
В
2624
В95-8

41.3
28.1
28.2
2623
Сао
46.1
pSG5
34.1
2624

63 кДа

Рис. 7. Активация сигнальных путей NF-kB (А) и АР-1 (Б) при экспрессии низкодивергентных и контрольных вариантов LMP1 (В).
4.2. Анализ активации iNOS и определение уровня NO при постоянной экспрессии низкодивергентных вариантов LMP1
в клетках крысиных фибробластов Rat-1
Известно, что в индукции iNOS белком LMP1 ключевую роль играют транскрипционные факторы NF-κB и STAT3. Более того, ранее показанные различия в активации iNOS и уровнях внутриклеточной генерации NO между отдельными вариантами LMP1 позволяют нам предположить влияние мутаций LMP1 на этот процесс [Yu et al., 2002, Дидук и др., 2008]. Для определения значения ограниченного числа замен исследуемых вариантов LMP1 в индукции образования окиси азота, мы анализировали линии Rat-1, постоянно экспрессирующие изучаемые низкодивергентные онкобелки LMP1, а также контрольные варианты LMP1-B95-8 и LMP1-Cao.
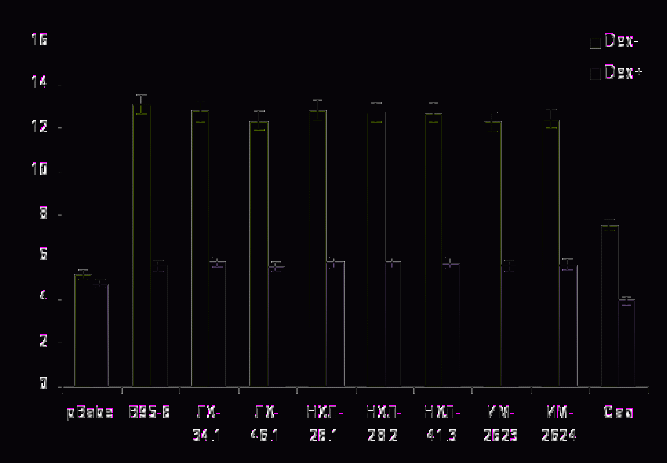
Рис. 8. Накопление нитратов/нитритов в культуральной среде клеточных линий Rat-1, экспрессирующих исследуемые варианты LMP1 (+/- дексаметазон).
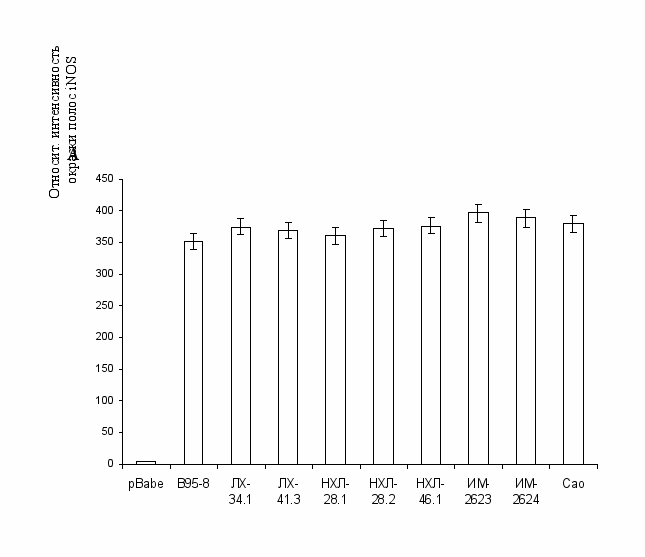

Рис. 9. Индукция iNOS при экспрессии вариантов LMP1: А – денситометрический анализ активации iNOS клеточными линиями Rat-1, экспрессирующими исследуемые варианты LMP1. Б – вестерн-блот-анализ с использованием антител, специфичных к LMP1 (S12), iNOS (NOS-20) и β-катенину. β-катенин использовали в качестве контроля нанесения.
С этой целью в 3-дневной культуральной среде исследуемых клеток Rat-1 измеряли концентрацию конечных продуктов нитратов и нитритов. Как видно на рис.8, количество выделяемого в культуральную среду NO клетками, экспрессирующими исследуемые варианты LMP1 российского происхождения, было несколько ниже по сравнению с прототипным вариантом LMP1-В95-8, но не снижалось до уровня NO, выделяемого клетками, экспрессирующими высокотуморогенный вариант LMP1-Cao. Так, при экспрессии вариантов ИМ-2623, ИМ-2624 и ЛХ-46.1 наблюдалось снижение уровня накопления NO на 4,6-5,4% (р ≥ 0,05) по сравнению с экспрессией прототипного LMP1-B95.8, хотя при экспрессии других низкодивергентных вариантов (ЛХ-34.1, НЛХ-28.1, НЛХ-28.2, НЛХ-41.3) это различие было не столь значительным.
Для подтверждения участия в индукции внутриклеточного синтеза NO индуцибильной формы NO-синтазы (iNOS), нами использовались специфические антитела к iNOS (рис. 9), а также дексаметазон (Dex), который является ингибитором этой формы фермента. Обработка в течение трех дней этим ингибитором клеток, экспрессирующих разные варианты LMP1, привела к снижению уровня синтеза NO: в случае прототипного варианта – на 60%, при экспрессии низкодивергентных вариантов – на 52-55%, а в случае Cao – на 46% (р < 0,001, рис. 8). Исходя из полученных результатов, можно сделать вывод об определенном влиянии даже небольшого набора мутаций (характерных для преобладающих в России вариантов LMP1) на уровень активации iNOS и внутриклеточное образование NO и, соответственно, на степень туморогенности вариантов LMP1.
ВЫВОДЫ
- Филогенетический анализ секвенированных полноразмерных вариантов LMP1, изолированных от российских больных лимфомой Ходжкина (ЛХ), неходжкинскими лимфомами (НХЛ) и инфекционным мононуклеозом (ИМ), выявил доминирование вариантов LMP1, относящихся к белкам низкодивергентной группы LMP1-B95.8b, характеризующейся минимальным набором мутаций.
- Получена коллекция клеточных линий на основе крысиных фибробластов Rat-1, постоянно экспрессирующих низкодивергентные варианты LMP1.
- При анализе морфологии полученных клеточных линий Rat-1, постоянно экспрессирующих варианты LMP1 российского происхождения, выявлена их способность к ингибированию контактного торможения и формированию колоний при росте без подложки.
- Показано, что экспрессия изучаемых вариантов LMP1 сопровождалась усилением активации транскрипционного фактора NF-κB и не оказывала влияния на активацию транскрипционного фактора АР-1.
- Выявлено, что все изучаемые низкодивергентные варианты LMP1, экспрессирующиеся в клетках Rat-1, вызывают активацию индуцибильной NO-синтазы (iNOS) и продукцию окиси азота (NO), при этом уровень накопления последнего оказался сходным с таковым, вызванным прототипным низкотрансформирующим вариантом LMP1-B95.8, но выше, чем у высокотуморогенного варианта LMP1-Cao.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- Павлиш О.А., Дидук С.В., Смирнова К.В., Щербак Л.Н., Гончарова Е.В., Шалгинских Н.А., Архипов В.В., Кичигина М.Ю., Степина В.Н., Белоусова Н.В., Османов Е.А., Яковлева Л.С., Гурцевич В.Э. Mутации гена LMP1 вируса Эпштейна-Барр у российских больных лимфоидной патологией и здоровых лиц. // Вопр. вирусол., - 2008. – №1, c. 10-16.
- Дидук С.В., Смирнова К.В., Павлиш О.А., Гурцевич В.Э. Роль функционально значимых мутаций гена LMP1 вируса Эпштейна-Барр в активации клеточных сигнальных путей.// Биохимия, - 2008, - №10, c. 1414-1421.
- Смирнова К.В., Дидук С.В., Гурцевич В.Э. Функциональный анализ вариантов латентного мембранного белка 1 (LMP1) вируса Эпштейна-Барр у больных лимфопролиферативными заболеваниями. // Биомедицинская химия, - 2009. - принята в печать.
- Смирнова К.В., Дидук С.В., Яковлева Л.С., Гончарова Е.В., Щербак Л.Н., Кичигина М.Ю., Михайлова Н.А., Степина В.Н., Огородникова Е.В., Османов Е.А., Павлиш О.А., Гурцевич В.Э.. Молекулярная эпидемиология гена LMP1 вируса Эпштейна-Барр: генетические варианты у больных опухолями лимфоидного и эпителиального происхождения в России. // Мол. Медицина, - 2010. - № 1, принята в печать.
- Дидук С.В., Смирнова К.В., Шалгинских Н.А. Сиквенсный анализ последовательностей гена LMP1 вируса Эпштейна-Барр у больных лимфопролиферативными заболеваниями в России. Материалы Всероссийской научно-практической конференции «Отечественные противоопухолевые препараты». // Российский биотерапевтический журнал, - 2007. - №1, с. 5.
- Гурцевич В.Э., Гончарова Е.В., Щербак Л.Н., Дидук С.В., Смирнова К.В., Шалгинских Н.А., Кичигина М.Ю., Степина В.Н., Яковлева Л.С., Павлиш О.А. Вирус Эпштейна-Барр (ВЭБ) как представитель В-лимфотропных герпесвирусов приматов: генетические варианты гена LMP1 в образцах ВЭБ российского происхождения. Материалы международной научной конференции «Фундаментальные и прикладные проблемы медицины и биологии в опытах на обезъянах», 2007, с. 170-182.
- Pavlish O.A., Diduk S.V., Smirnova K.V., Scherback L.N., Goncharova E.V., Shalginskykh N.A., Kichigina M.Yu., Arkhipov V.V., Belousova N.V., Stepina V.N., Osmanov E.A., Yakovleva L.S., Gurtsevitch V.E. Epstein-Barr virus LMP1 gene variants in russian patients with limphoproliferative disorders. Abstracts of 16-th International Conference “AIDS, Cancer and Public Health”. // Russian journal AIDS, сancer and public health, - 2007. - V. 11, № 1, p. 123.
- Diduk S.V., Smirnova K.V., Pavlish O.A., Gurtsevitch V.E. Conservative mutations of latent membrane protein 1 (G212S/S350A/S366T) in Epstein-Barr virus-mediated activation signal pathways. Abstracts of the Third European Congress of Virology, 2007, p. 244.
- Diduk S.V., Smirnova K.V., Gurtsevitch V.E. The significance of some mutations of EBV LMP1 gene for strengthening its oncogenic potential. Abstracts of the 8th John Humphrey Advanced Summer Program In Immunology, 2007, p.52.
- Gurtsevitch V., Smirnova K., Diduk S., Goncharova E., Scherback L., Stepina V., Yakovleva L. Molecular epidemiology of the Epstein-Barr virus (EBV) LMP1 genetic variants of Russian origin. Abstracts of 5th Russian-German Conference “Human herpesvirus infection”, 2008, p. 10.
- Дидук С.В., Смирнова К.В., Гурцевич В.Э. Влияние мутаций гена LMP1 на внутриклеточную генерацию активных форм азота как важнейшего фактора в усилении трансформирующей активности вируса Эпштейна-Барр. Материалы Всероссийской научно-практической конференции «Отечественные противоопухолевые препараты». // Российский биотерапевтический журнал, - 2008. - №1, с. 9.
- Дидук С.В., Смирнова К.В., Гурцевич В.Э. Влияние мутаций области CTAR гена LMP1 герпесвируса Эпштейна-Барр (ВЭБ) на его трансформирующую активность. Тезисы 16-ой Международной конференции «СПИД, рак и общественное здоровье». // Русский журнал СПИД, рак и общественное здоровье, - 2008. - Т. 12, №2, с. 90.
- Diduk S.V., Smirnova K.V., Gurtsevitch V.E. Nitric oxide production in Rat-1 cells induced by Epstein-Barr virus LMP1 gene with mutations in canonical and cryptic HOS recognition sites. Abstracts of the 17th International Conference “AIDS, Cancer and Public Health”. // Russian journal AIDS, сancer and public health, - 2008. - V. 12, № 2, p. 42.
- Diduk S., Smirnova K., Gurtsevitch V. Activation of some signaling pathways in Rat-1 cells induced by Epstein-Barr virus LMP1 gene with mutations in HOS recognition sites. // European Journal of Cancer, Suppl., European Association for Cancer Research (EARC) 20 Meeting, - 2008. - Lyon, France, V. 6, № 9, p. 148.
- Gurtsevitch V., Smirnova K., Diduk S., Goncharova E., Scherback L., Stepina V., Yakovleva L. LMP1 gene variants of Russian origin from patients with EBV-associated pathologies and health individuals. // European Journal of Cancer, Suppl., European Association for Cancer Research (EARC) 20 Meeting, - 2008. - Lyon, France, V. 6, № 9, p. 50.
- Смирнова К.В., Дидук С.В., Гурцевич В.Э. Филогенетический анализ вариантов гена LMP1 вируса Эпштейна-Барр у больных лимфопролиферативными заболеваниями в России. Материалы VIII Всероссийская научно-практическая конференция с международным участием «Отечественные противоопухолевые препараты». // Российский биотерапевтический журнал, - 2009. - № 4, с. 11.
- Смирнова К.В., Дидук С.В., Гурцевич В.Э. Анализ активации ключевых транскрипционных факторов низкодивергированными вариантами гена LMP1 вируса Эпштейна-Барр, полученными от больных лимфопролиферативными заболеваниями в России. Анализ активации ключевых транскрипционных факторов низкодивергированными вариантами гена LMP1 вируса Эпштейна-Барр, полученными от больных лимфопролиферативными заболеваниями в России. Материалы Российской конференции по фундаментальной онкологии. // Вопросы онкологии, - 2009. - № 4, с.15.