
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Естественная и антропогенная динамика лиственничных лесов криолитозоны (на примере Якутии)
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАОбщее представление о характере роста и развития древесной растительности на вырубках можно получить из рис. 11. Сравнивая кривые, составленные для вырубок в разных типах леса, можно выделить два направления (стратегии) в обеспечении успешности лесовозобновления. Первое характерно для вырубок в лиственничниках свежих и средневлажных условий произрастания, наиболее широко представленных в северной части среднетаежной Якутии. Успешность лесовозобновления в них определяется обилием подроста. Второе направление отмечается для вырубок в лиственничниках с елью и кедром бруснично-зеленомошных, широко представленных в Юго-Западной Якутии. Успешность лесовозобновления в них определяется большей интенсивностью роста.
В целом вырубки, возникшие в результате промышленных лесозаготовок, отличаются значительно большим количеством подроста, более равномерным его размещением и, соответственно, более надежным лесовозобновлением, чем вырубки, разработанные с использованием конной трелевки.
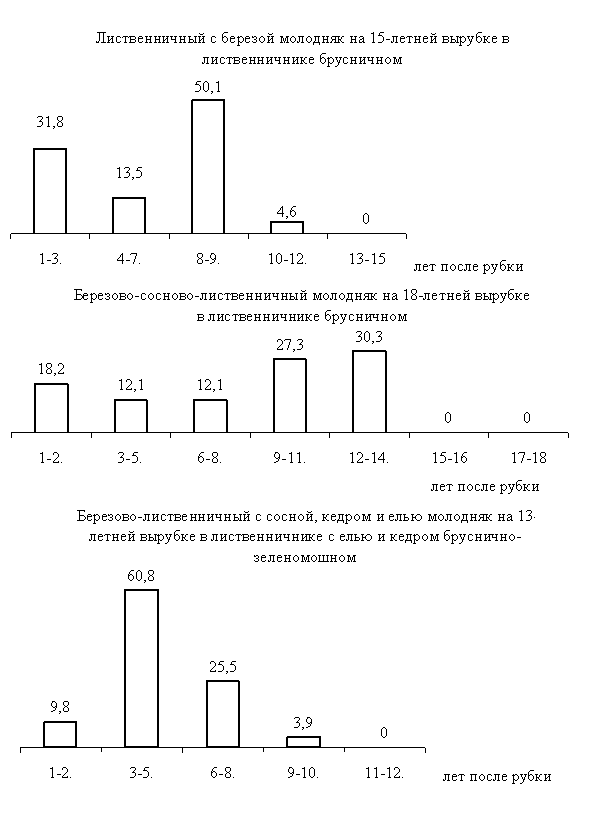
Рис. 10. Этапы появления семосева древесных пород на вырубках,
в % от общего числа самосева
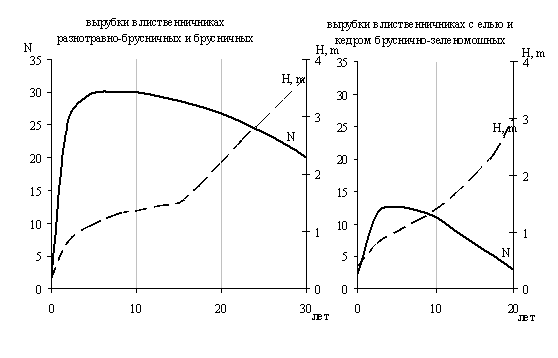
Рис. 11. Динамика численности (N, тыс. экз./га) и средней высоты подроста (H, m)
на вырубках в лиственничных лесах
5.1.5. Динамика растительности в шелкопрядниках. Изменение обстановки после гибели древостоя в шелкопряднике ведет к изменению состава и структуры растительности.
В первый год различия по составу не столь велики, однако заметно изменяются количественные характеристики стенобионтных лесных видов, которые постепенно снижают свою численность и проективное покрытие и к 3-му году после нарушения полностью выпадают из ценоза. Некоторые лесные виды, характеризующиеся более широким экологическим ареалом, снижают свое обилие постепенно, но полностью не исчезают или исчезают на более поздних стадиях (Vaccinium vitis-idaea, Linnaea borealis, Limnas stelleri). Часть видов в первые годы даже увеличивают свое обилие (Artemisiatanacetifolia, Moerhingialaterifolia, Poasibirica, Vicia cracca, Fragaria orientalis). Начиная с 1-го года, происходит процесс олуговения, приобретающий к 3-4-му году вполне ощутимые очертания. Начинают доминировать луговые и рудеральные виды.
Самосев лиственницы и березы на части площадей шелкопрядников на 3-1 год после гибели древостоя отсутствовал, лишь в отдельных случаях отмечались отдельные всходы лиственницы и активизация порослевой березы. Даже на 9-й год после нарушения в ряде случаев имелись лишь единичные экземпляры лиственниц. В связи с отсутствием на части площадей шелкопрядников самосева березы и лиственницы, лесовосстановительный процесс затягивается на многие годы, и после 10 лет после гибели леса шелкопрядники представаляют собой симбиоз погибщего леса (сухостой) и луговой растительности.
5.1.6.Общая схема экзогенных сукцессий лиственничных лесов и закономерности формирования климаксовых сообществ. При большом количестве сохраненного предварительного подроста или массовом появлении самосева лиственницы лесовосстановление на вырубках и гарях в лиственичниках в большинстве случаев не вызывает сомнения. Если же исключается возможность непросредственного (впервые 2-5 лет) возобновления лиственницы, условия для появления самосева не могут возникнуть и в последующем. Таким образом, при достаточном количестве лиственничного подроста сукцессионное развитие растительности протекает без смены пород. Образующийся обычно березовый (с участием высоких кустарников) полог следует рассматривать лишь в качестве восстановительной стадии, так как он не устойчив, кратковременен и поэтому не может свидетельствовать о смене пород. Формирование березового леса, устойчивого, по крайней мере, в течение одного поколения березы (лишь в этом случае следует говорить об истинной смене пород), происходит довольно редко.Направление и ход лесообразовательного процесса резко изменяются при возможных проявлениях термокарста. Вторичная сукцессия в этом случае представляет собой не обычную демутацию, а квазипервичную. При этом могут развиваться как более влажные типы лиственничников, так и ерниковые формации, травяные болота или провальные озера.
Обобщая разнообразие имеющихся сведений о растительном покрове, составлена принципиальная схема изменения растительного покрова в связи с экзогенными причинами в лиственничниках брусничной группы (рис. 12).
При этом лиственничники нельзя рассматривать в качестве классических климаксовых формаций. Если в лесах Юго-Западной и Южной Якутии климаксовыми лесами следует считать темнохвойные леса (еловые, кедровые и пихтовые), то в Северной и Центральной Якутии, частично в Западной и Юго-Восточной Якутии климаксовых лесов (в полном понимании этого слова) нет. Здесь безраздельно господствует лиственничная тайга.
Лиственница, являясь светлохвойной породой, типичным эксплерентом (в начале сукцессионного процесса) и патиентом на последующих стадиях, не может формировать климаксовые сообщества. Под пологом в условиях отсутствия внешнего возмущающего фактора лиственница не возобновляется естественным образом (Шурдук, Исаев, 1991). Господство же ее объясняется приспособленностью лиственницы к произрастанию в условиях близкого залегания многолетней мерзлоты и биологически обусловленной зависимостью лесовозобновления лиственничной тайги от лесных пожаров. Лиственничные леса в связи с этим и здесь можно считать квази- или ложноклимаксовыми.
аа 5.2. Функциональные закономерности формирования лиственничных лесов криолитозоны
5.2.1. Рост, развитие и фитомасса лиственничных лесов редколесий у северной границы леса. Характеристику притундровых лесов приводим на примере растительности острова Тит-Ары в низовьях р. Лены. После 1901 г., когда остров посетил А.К. Каяндер, более или менее регулярные исследования лиственничных редколесий на острове Тит-Ары начались в 1955 г. Результатами исследований явился ряд работ (Б.А. Тихомиров, В.С. Штепа, 1956; Т.Г. Полозова, 1961; И.П. Щербаков, 1965; В.Н. Андреев, 1975; Растительный и животный мир дельты р. Лены, 1985; В.И. Перфильева, Л.В. Тетерина, Н.С. Карпов, 1991). К сожалению, материалы о структуре древостоев в основном носили неполный, отрывочный характер. В связи с этим не все параметры поддаются оценке их изменения (табл. 16). Густота стояния деревьев начиная с 1955 г. постепенно снизилась с 1,5 до 0,7 тыс. экз./га. По показателям высоты (3-5 м) деревья практически достигли, а по толщине пока еще значительно уступают показателям дорубочного насаждения.
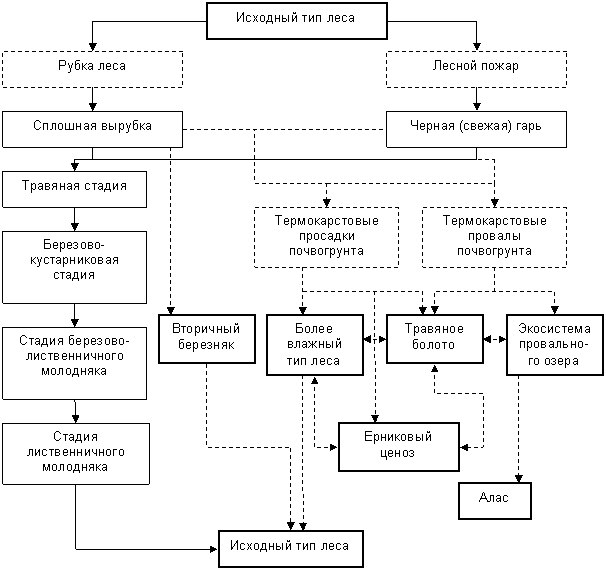

Рис. 12. Принципиальная схема изменения растительного покрова на гарях и вырубках в лиственничниках брусничной группы
1 - исходный тип леса и производные фитоценозы;
2 - стадии сукцессионого развития
3 - основные факторы, влияющие на ход сукцессий;
4 - наиболее типичные направления сукцессий;
5 - возможные направления сукцессий
аТаблица 16
Вековая динамика основных таксационных показателей лиственничного
редколесья пушицево-мохового на острове Тит-Ары. Булунский улус
Годы наблюдений |
число стволов, тыс.экз./га |
господствующая высота, м |
господствующий диаметр, см |
1901 |
нет данных |
3-5 |
нет данных |
1942* |
нет данных |
до 5 |
10-15 |
1942-1943 |
был вырублен |
||
1955 |
1,5 |
нет данных |
нет данных |
1962 |
1,5 |
нет данных |
1-4 |
1984 |
1,5 |
0,3-1,6 (2,5) |
нет данных |
1988 |
нет данных |
0,3-3,0 |
2(4) |
1989 |
0,8 |
1,9-2,8 |
1,5 (3,5) |
2009 |
0,7 |
2,15 (4, 5) |
2,06 (4,7) |
Общая надземная фитомасса древостоя в разных условиях составляет 557,1-3383,9 кг/га (табл. 17). Причем практически во всех типах леса доля хвоевой части высока, что говорит об относительной молодости древостоев и их низкой продуктивности.
Таблица 17
Надземная фитомасса древостоев различных типов лиственничных редколесий острова Тит-Ары, Булунский район РС (Я)
№ пр. пл. |
Типы лиственничных редин и редколесий |
Надземная фитомасса, т/га |
||
древесина |
хвоя |
общая |
||
ТА-5 |
.р. кустарниково-моховое |
0,603 |
0,065 |
0,668 |
ТА-6 |
.р. кассиопово-моховое |
1,723 |
0,183 |
1,906 |
ТА-9 |
.р. голуб.-багульн.-моховая |
3,089 |
0,295 |
3,384 |
ТА-10 |
.р. багульниковое |
0,880 |
0,083 |
0,963 |
ТА-13 |
.р. пушицево-моховое |
0,503 |
0,054 |
0,557 |
Х-4 |
.р. багульн.-лишайн.-моховое |
0,500 |
0,059 |
0,559 |
5.2.2. Рост, развитие и фитомасса лиственничных редколесий у высотной границы леса в горах.Рассмотрены процессы роста и развития лиственничных древостоев в условиях высотной границы леса (хребет Черского, Оймяконский улус). Высотная граница леса расположена на высоте 1000-1200 м над ур.м. Выявлено, что фитомасса у лиственничных редколесий у высотной границы леса в разы выше (табл. 18), чем фитомасса у широтной границы.
Таблица 18
Фитомасса древостоя лиственничного редколесья ерникового багульниково-мохового. Оймяконский улус
Компоненты древостоя |
Фитомасса |
|
т сух. в-ва / га |
% |
|
Надземная часть, в .ч. |
7,55 |
61,5 |
Ствол |
6,42 |
9,0 |
Ветки |
0,93 |
1,9 |
Хвоя |
0,20 |
72,4 |
Подземная часть (скелетные корни) |
2,88 |
27,6 |
Общая фитомасса |
10,43 |
100,0 |
5.2.3. Рост, развитие и динамика фитомассы среднетаежных лиственничных лесов. За период исследований (1996-2001 г.) в подзоне средней тайги в пределах Центральной Якутии изучалась фитомасса лиственничников брусничной группы. Всего было заложено 38 пробных площади, в том числе 17 - в лиственничниках брусничных. Для анализа привлечены также материалы по 7 пробным площадям других авторов (Поздняков и др., 1969; Kanazawa et al., 1994).
Лесные участки лиственничников брусничной группы представляли разновозрастные древостои, в связи с чем появилась возможность выстроить 250-летний динамический ряд таксационных показателей и надземной фитомассы (рис. 13).
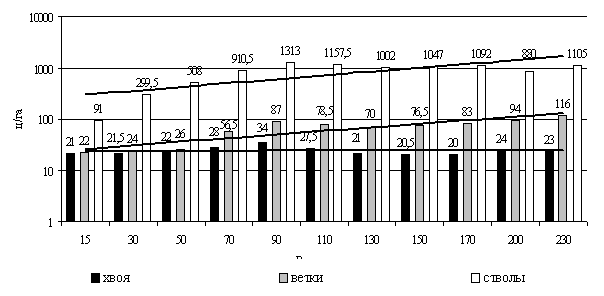
Рис. 13. Динамика фитомассы брусничных лиственничников в Центральной Якутии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии