
Книга вторая Дж. Эдвард Морган-мл. Мэгид С. Михаил Перевод с английского
| Вид материала | Книга |
- Книга первая Дж. Эдвард Морган-мл. Мэгид С. Михаил Перевод с английского, 10010.77kb.
- А. Конан-Дойль новоеоткровени е перевод с английского Йога Рàманантáты, 2314.23kb.
- Copyright Сергей Александровский, перевод с английского Email: navegante[a]rambler, 619.61kb.
- "книга непрестанности осириса " 177, 7373.41kb.
- Н. М. Макарова Перевод с английского и редакция, 4147.65kb.
- Трудового Красного Знамени гупп детская книга, 2911.61kb.
- Трудового Красного Знамени гупп детская книга, 2911.77kb.
- Уайнхолд Б., Уайнхолд Дж. У 67 Освобождение от созависимости / Перевод с английского, 11462.2kb.
- Малиновской Софьи Борисовны Специальность: журналистика Специализация: художественный, 969.08kb.
- Духовные истины в психических явлениях перевод с английского 3-е издание Москва «Философская, 1557.75kb.
Петля Генле состоит из нисходящей и восходящей частей. Тонкий сегмент нисходящей части является продолжением проксимального канальца и опускается из коркового вещества почки в мозговое. В мозговом веществе тонкий сегмент загибается, делая U-образный разворот, и поднимается в корковое вещество уже как восходящая часть петли Генле. В восходящей части выделяют различающиеся в функциональном отношении тонкий сегмент, толстый медуллярный сегмент и толстый кортикальный сегмент (рис. 31-1). У нефронов, клубочки которых расположенные вблизи мозго-
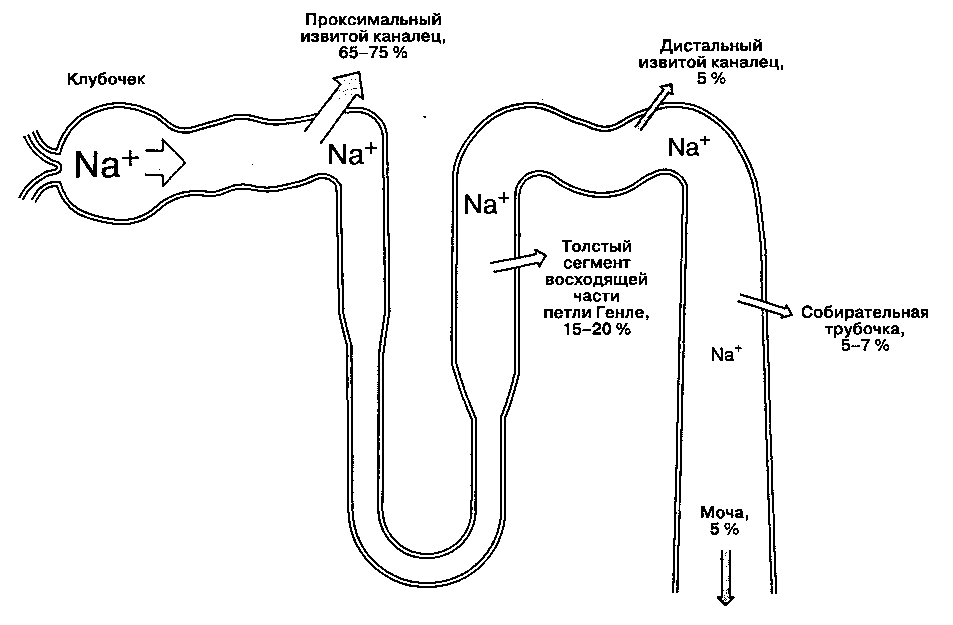
Рис. 31-2. Реабсорбция натрия в нефроне. Указанное количество реабсорбированного натрия в каждом отделе нефро-на соответствует проценту от профильтровавшейся натриевой нагрузки. (Из: Cogan M. G. Fluid and Electrolytes: Physiology and Pathophysiology, 14th ed. Appleton & Lange, 1991; с разрешения.)
вого вещества (юкстамедуллярные нефроны), петля Генле более длинная, чем у нефронов, клубочки которых лежат ближе к наружной поверхности почки (корковые нефроны). У корковых нефронов с короткой петлей отсутствует тонкий сегмент восходящей части. Корковых нефронов в 7 раз больше, чем юкстамедуллярных. Петля Генле поддерживает гипертоничность интерстициальной жидкости мозгового вещества, а также опосредованно обеспечивает процесс концентрирования мочи в собирательных трубочках.
В норме до 25-35 % фильтрата, образующегося в капсуле Боумена, достигает петли Генле. В ней реабсорбируется 15-20% фильтруемого натрия. За исключением толстого сегмента восходящей части, реабсорбция растворенных веществ и воды в петле Генле происходит пассивно — по градиенту концентрации и осмотическому градиенту соответственно. В толстом сегменте восходящей части Na" и СГ реабсорбируются в большей степени, чем вода; более того, в этой части нефрона реабсорбция Na" непосредственно сопряжена с реабсорбцией K+ и СГ (рис. 31-4), и концентрация СГ в канальцевой жидкости является фактором, ограничивающим
скорость реабсорбции. Активная реабсорбция Na+ осуществляется Ка"УК+-зависимой АТФ-азой капиллярной поверхности эпителиальных клеток.
В отличие от нисходящей части и тонкого сегмента восходящей части петли Генле, толстый сегмент восходящей части непроницаем для воды. Поэтому оттекающая из петли Генле канальцевая жидкость гипотонична (100-200 мОсм/л), а окружающая петлю Генле интерстициальная жидкость гипертонична. Механизм противоточного умножения работает таким образом, что гипертонич-ностъ каналъцевой жидкости и окружающего uh-терстиция значительно нарастает по мере углубления в мозговое вещество почки (рис. 31-5). Концентрация мочевины в мозговом веществе становится высокой, что существенно влияет на его гипертоничность. Структуры механизма противоточного умножения включают петлю Генле, кортикальные и медуллярные собирательные трубочки и сопровождающие их капилляры (vasa recta),
Толстый сегмент восходящей части петли Генле играет важную роль в реабсорбции Ca2+ и Mg2+. В этом участке нефрона паратиреоидный гормон может увеличивать реабсорбцию кальция.
^ Дистальный извитой каналец
В дистальный извитой каналец поступает гипото-ничная жидкость из петли Генле. Как правило, состав и объем канальцевой жидкости в этом отделе нефрона изменяется значительно. В отличие от более проксимальных отделов нефрона, клетки эпителия дистальных канальцев плотно прилежат друг к другу, что делает их относительно непроницаемыми для воды и натрия, за счет чего поддерживаются градиенты концентраций, сформированные петлей Генле. В дистальных канальцах реабсорбируется 5 % фильтруемого в клубочках натрия. Реабсорбция натрия обеспечивается за счет функционирования КауК+-зависимой АТФ-азы на обращенной к капилляру поверхности клеток, а на люминальной мембране осуществляется Na^/СГ-переносчиком. Реабсорбция натрия в дистальном канальце прямо пропорциональна его поступлению. Дистальные канальцы — основное место реабсорбции кальция; этот процесс регулируется паратиреоидным гормоном и витамином D.
Наиболее дистальный отдел канальца называют соединяющим сегментом. В этом участке, как и в более проксимальных, происходит опосредованная гормонами реабсорбция кальция, но в отличие от них здесь имеет место еще и регулируемая альдостероном реабсорбция натрия.
^ Собирательная трубочка
В собирательной трубочке реабсорбируется 5-7 % фильтруемого в клубочках натрия, в ней выделяют кортикальную и медуллярную части.
^ А. Кортикальная часть собирательной трубочки. Этот сегмент нефрона состоит из двух типов клеток: 1) главных клеток, секретирующих калий и участвующих в опосредованной альдостероном реабсорбции натрия; 2) вставочных клеток, ответственных за регуляцию кислотно-основного равновесия. Главные клетки реабсорбируют натрий с помощью электрогенной помпы, поэтому для поддержания электронейтральности должна происходить либо реабсорбция СГ, либо секреция K+.

Рис. 31-3. Реабсорбция фосфатов, глюкозы, аминокислот и электролитов в проксимальном извитом канальце. Отметим, что №+/Кт-зависимая АТФ-аза, поддерживая низкую концентрацию натрия внутри клеток канальце-вого эпителия, обеспечивает энергетические потребности для реабсорбции катионов
Рис. 31-4. Реабсорбция натрия и хлора в толстом сегменте восходящей части петли Генле. Белок-переносчик (расположенный на люминальной поверхности клетки петли Генле) функционирует, только если заняты все четыре места связывания. Концентрация хлора в канальцевой жидкости является фактором, ограничивающем скорость реабсорбции
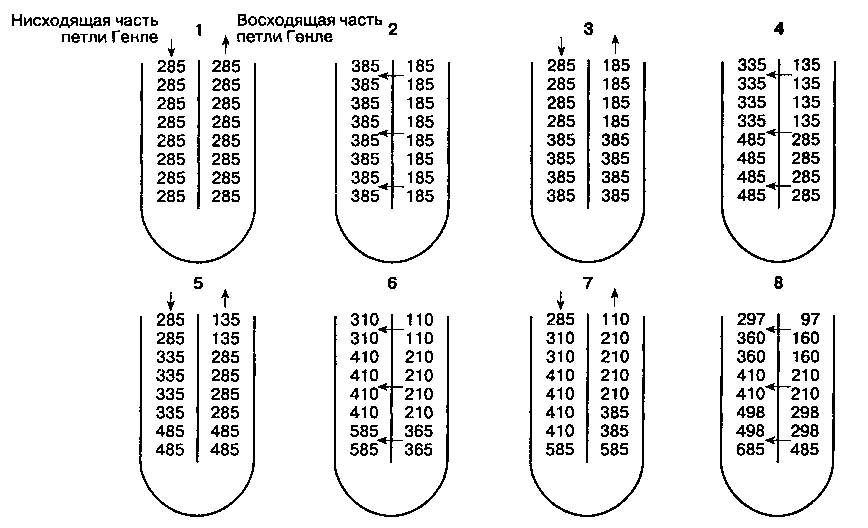
Рис. 31-5. Механизм противоточного умножения, обу слов лен ный различиями в проницаемости и трансмембранном транспорте, которые существуют между нисходящей и восходящей частями петли Генле. Нисходящая часть и тонкий сегмент восходящей части петли Генле проницаемы для воды, Na+, Cl и мочевины. Толстый сегмент восходящей части петли Генле непроницаем для воды и мочевины, в нем активно реабсорбируется Na+ и Cl . Эти факторы приводят к возникновению осмотического градиента. На рисунке представлен осмотический градиент между нисходящей и восходящей частями петли Генле, постепенно увеличивающийся от О до 200 мОсм/кг. Заметим, что в петле Генле по ходу тока канальцевой жидкости градиент не изменяется, в то время как осмоляльность увеличивается по мере приближения к точке перегиба. (По: Pitts R. F. Physiology of the Kidney and Body Fluids, 3rd ed. Year Book, 1974.)
Увеличение концентрации внутриклеточного калия способствует его секреции. В данном отделе нефрона альдостерон повышает активность NaV К+-зависимой АТФ-азы, увеличивая количество открытых K+- и Na^-каналов в люминальной мембране. Альдостерон также повышает активность ГГ-секретирующей АТФ-азы на люминальной поверхности вставочных клеток (рис. 31-6). Кроме того, во вставочных клетках люминальная Ю/Н+-зависимая АТФ-аза реабсорбирует K+ и секрети-рует ГГ. Некоторые вставочные клетки способны секретировать бикарбонат при большой щелочной нагрузке.
^ Б. Медуллярная часть собирательной трубочки — основная мишень антидиуретического гормона (АДГ, синоним — аргинин-вазопрессин). АД Г активирует аденилатциклазу через У2-рецеп-торы (активация V1-рецепторов повышает сосудистое сопротивление за счет усиления метаболизма фосфатидилинозитола). Проницаемость люминальной мембраны для воды полностью зависит от присутствия АДГ (гл. 28). Дегидратация приводит к повышению секреции АДГ, в результате
действия которого люминальная мембрана становится проницаемой для воды. По осмотическому градиенту вода удаляется из канальцевой жидкости, и образуется концентрированная моча (до 1400 мОсм/л). Напротив, адекватная гидратация подавляет секрецию АДГ; в этом случае осмоляльность канальцевой жидкости в медуллярной части собирательных трубочек не изменяется, и она остается гипотоничной (100-200 мОсм/л). В стенке медуллярной части собирательных трубочек содержатся основные и вставочные клетки, причем последние преобладают. Кроме того, эта часть нефрона отвечает за подкисление мочи; образующиеся в этих клетках ионы водорода выделяются в виде титруемых кислот (фосфаты) и ионов аммония (гл. 30). Предсердный натрийуретический пептид способен снижать реабсорбцию Na" в собирательных трубочках.
^ В. Роль собирательных трубочек в поддержании гипертоничности мозгового вещества. Кортикальная часть собирательных трубочек полностью проницаема для мочевины, тогда как медуллярная часть в норме для нее непроницаема. Этим разли-
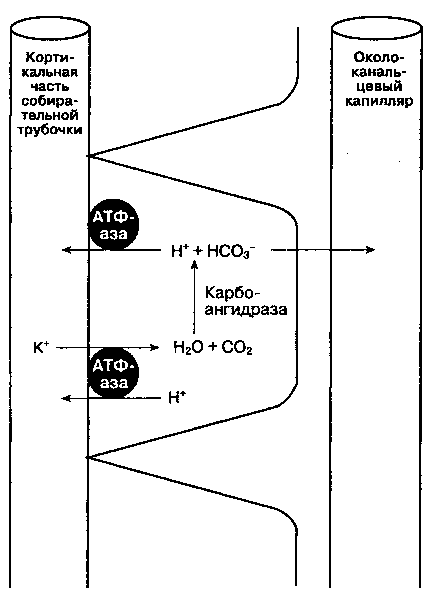
Рис. 31-6. Кортикальная часть собирательной трубочки: секреция ионов водорода, реабсорбция бикарбоната и калия
чием в проницаемости практически наполовину обусловлена гипертоничность интерстициальной жидкости мозгового вещества. В присутствии АДГ значительно увеличивается проницаемость для мочевины наиболее дистальных отделов медуллярной части собирательных трубочек. Таким образом, при секреции АДГ вода покидает собирательные трубочки, и моча становится более концентрированной. Затем мочевина диффундирует в интерстициальную жидкость мозгового вещества, увеличивая ее осмоляльность.
^ Юкстагломерулярный аппарат
Этот небольшой орган, имеющийся в каждом неф-роне, состоит из специализированного сегмента приносящей артериолы, в стенке которой содержатся юкстагломерулярные клетки, и конечной части толстого сегмента восходящей части петли Генле — плотного пятна (macula densa\ рис. 31-7). Юкстагломерулярные клетки содержат фермент ренин и иннервируются симпатическими нервными волокнами. Факторы, влияющие на высвобождение ренина: 1) стимуляция fii-адренорецепто-ров; 2) давление в приносящей артериоле (гл. 28); и 3) концентрация ионов хлора в жидкости,
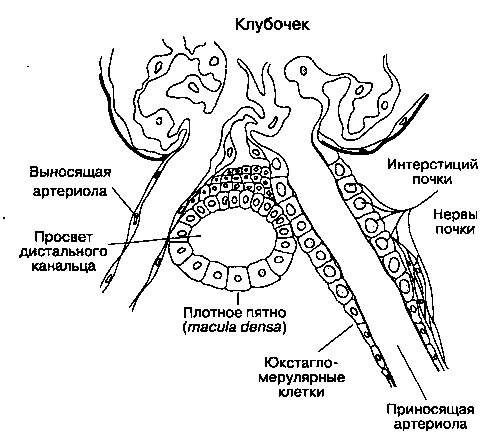
Рис. 31-7. Юкстагломерулярный аппарат. (С изменениями. Из: Ganong W. F. Review of Medical Physiology, 14th ed. Appleton & Lange, 1989.)
протекающей через плотное пятно. Высвобождаемый в кровоток ренин воздействует на продуцируемый печенью белок — ангиотензиноген, в результате чего образуется ангиотензин I. Под воздействием ангиотензинпревращающего фермента (АПФ) физиологически инертный декапептид ангиотензин I быстро превращается в октапептид ангиотензин II. Процесс идет главным образом в сосудах легких. Ангиотензин II играет важную роль в регуляции АД (гл. 19) и секреции альдостерона (гл. 28). В клетках прокси-мальных канальцев содержатся АПФ, а также рецепторы ангиотензина IL Кроме того, внутрипо-чечное образование ангиотензина II увеличивает реабсорбцию натрия в проксимальных канальцах. Вне почек ренин и ангиотензин II вырабатываются в эндотелии сосудов, в надпочечниках и в ткани головного мозга.
^ Почечное кровообращение
Функция почек находится в тесной зависимости от кровотока в них. Почки — это единственный орган, в котором потребление кислорода зависит от кровотока; в других органах такой зависимости нет. На почечный кровоток (в обеих почках) приходится 20-25 % сердечного выброса. Кровь в почку поступает по почечной артерии, отходящей от аорты. В воротах почки почечная артерия разветвляется на междолевые артерии, от которых на границе коркового и мозгового вещества отходят дуговые
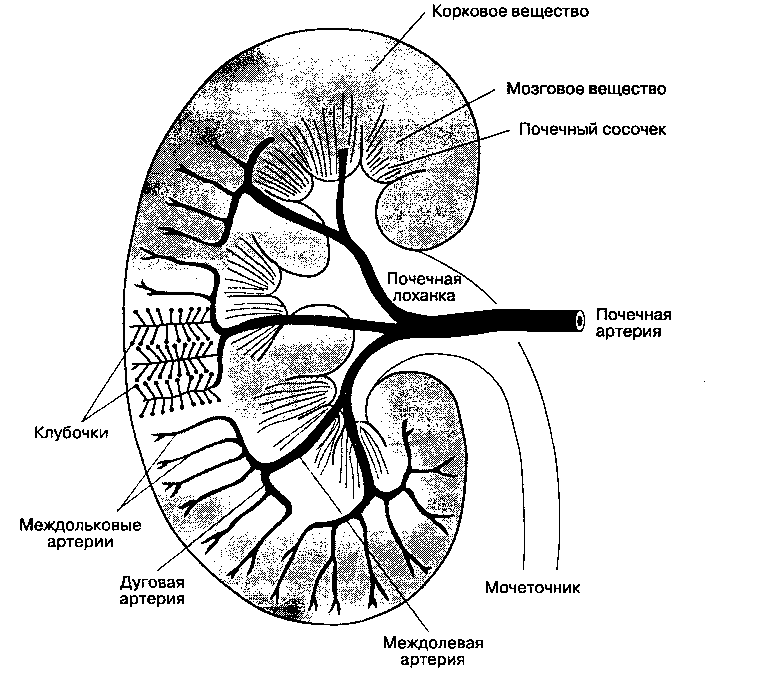
Рис. 31-8. Артерии почки. (С разрешения. Из: Leaf A., Cotran R. S. RenalPatophysiology. Oxford Univ. Press, 1976.)
артерии (рис. 31-8). Дуговые артерии делятся на междольковые артерии, которые отдают к каждому клубочку по одной приносящей артериоле. От каждого клубочка отходит единственная выносящая артериола, по которой осуществляется отток крови. В свою очередь выносящие артериолы распадаются на капилляры, оплетающие почечные канальцы. В отличие от капилляров клубочков, где происходит фильтрация, в околоканальцевых капиллярах осуществляется реабсорбция. Венулы, собирающие кровь из околоканальцевого капиллярного сплетения, через почечную вену возвращают кровь в нижнюю полую вену.
^ Почечный кровоток и клубочковая фильтрация
Клиренс
Концепция клиренса часто используется при оценке величины почечного кровотока (ПК) и скорости клубочковой фильтрации (СКФ). Почечный
клиренс вещества определяют как объем крови, полностью освобождаемый от этого вещества за единицу времени (обычно за 1 мин).
^ Почечный кровоток
Почечный плазмоток (ППТ) обычно определяют по клиренсу пара-аминогиппуровой кислоты (ПАГ). Если концентрация ПАГ в плазме низка, то справедливо допущение, что в течение одного пассажа через почки ПАГ полностью исчезает из плазмы за счет фильтрации и секреции. Следовательно,
ППТ = Клиренс ПАГ =
= ([ПАГ]моча /[ПАГ]плазма) х Диурез,
где [ПАГ]мома — это концентрация ПАГ в моче, а [ПАГ]ШШМа — это концентрация ПАГ в плазме. Если известен гематокрит, то
Почечный кровоток = ППТ/(1 - гематокрит).
В норме почечный плазмоток равен 660 мл/мин, почечный кровоток — 1200 мл/мин.
^ Скорость клубочковой фильтрации
Скорость клубочковой фильтрации в норме составляет около 20 % почечного плазмотока. Клиренс инулина (полисахарид фруктозы), который полностью фильтруется в клубочках, но не секре-тируется и не подвергается реабсорбции, адекватно отражает СКФ. В норме СКФ составляет 120 ± 25 мл/мин у мужчин и 95 ± 20 мл/мин у женщин.
По сравнению с клиренсом инулина клиренс креатинина отражает СКФ менее точно, но именно его используют на практике ввиду большего удобства расчета (гл. 32). При расчете по клиренсу креатинина СКФ обычно завышается, потому что незначительная часть креатинина секретируется печеными канальцами. Креатинин представляет собой продукт метаболизма фосфокреатина в мышцах. Клиренс креатинина рассчитывают еле-" дующим образом:
[Креатинин]м х Диурез
Клиренскреатинина= —:———————————— ,
[Креатинин]п
где [Креатинин]м — концентрация креатинина в моче, [КреатининJ11 — концентрация креатинина в плазме.
Отношение СКФ к ГШТ называется фракцией фильтрации (ФФ). В норме фракция фильтрации составляет 20 %. СКФ зависит от тонуса приносящих и выносящих артериол. Расширение приносящих или сужение выносящих артериол позволяют увеличить ФФ и поддержать СКФ даже при снижении почечного плазмотока. Изменение тонуса приносящих артериол обеспечивает постоянство СКФ даже при значительных колебаниях АД.
^ Механизмы регуляции
Выделяют ауторегуляшпо, канальцево-клубочко-вый баланс, гормональную и нервную регуляцию почечного кровотока. Эти механизмы регуляции взаимодействуют друг с другом.
^ А. Ауторегуляция. Благодаря ауторегуляции почечный кровоток (и СКФ) остается постоянным при изменении среднего АД от 80 до 180 мм рт, ст. Принцип ауторегуляции: при росте АД приносящие артериолы сужаются, при снижении АД — расширяются. При падении АД ниже 70 мм рт. ст. почечный кровоток снижается. Полагают, что изменение АД вызывает внутреннюю миогенную реакцию артериол, хотя точный механизм неизвестен. Вне рамок ауторегуляции почечный кровоток становится зависимым от АД. Клубочковая фильтрация прекращается при уменьшении среднего АД ниже 40-50 мм рт. ст.
^ Б. Канальцево-клубочковый баланс и обратная связь. Изменение скорости тока канальцевой жидкости влияет на СКФ: увеличение скорости тока канальцевой жидкости приводит к снижению СКФ, тогда как снижение, наоборот, способствует увеличению СКФ. Канальцево-клубочковая обратная связь, вероятно, играет важную роль в обеспечении постоянства СКФ в широком диапазоне клубочко-вого перфузионного давления. Хотя механизм этого феномена изучен недостаточно, установлено, что macula densa осуществляет канальцево-клубочко-вую обратную связь, вызывая рефлекторное изменение тонуса приносящей артериолы и, возможно, проницаемости капилляров клубочка. Вероятно, что ангиотензин II играет в нем посредническую роль. Местное высвобождение аденозина (которое происходит в ответ на увеличение ОЦК) способно подавлять секрецию ренина и расширять приносящие артериолы. Феномен прессорного натрийуре-за, т. е. снижения реабсорбции натрия при увеличении АД, также отражает канальцево-клубочковую обратную связь.
^ В. Гормональная регуляция. Повышение давления в приносящих артериолах стимулирует высвобождение ренина и образование ангиотензина II, Ангиотензин II вызывает генерализованную артериальную вазоконстрикцию и вторичное снижение почечного кровотока. Вазоконстрикция происходит как в приносящих, так и в выносящих артериолах, но диаметр последних меньше, поэтому их сопротивление увеличивается в большей степени, и СКФ практически не изменяется. Катехоламины, выделяемые надпочечниками (адреналин, норадрена-лин), повышают тонус приносящих артериол, но СКФ снижается минимально вследствие сопутствующего (опосредованного ими) высвобождения ренина и образования ангиотензина IL Относительная стабильность СКФ при увеличении секреции альдостерона или катехоламинов достигается за счет ангиотензининдуцируемого синтеза проста-гландинов и блокируется ингибиторами синтеза простагландинов (нестероидные противовоспалительные средства).
Напротив, дофамин в низких дозах расширяет приносящие и выносящие артериолы. Дофамин, образующийся в проксимальных канальцах, а также высвобождаемый нервными окончаниями, снижает реабсорбцию Na+ в проксимальных канальцах. Предсердный натрийуретический пептид расширяет приносящие артериолы, но сужает выносящие, существенно повышая СКФ (гл. 28). Кроме того, предсердный натрийуретический пептид уменьшает реабсорбцию Na+ в собирательных трубочках. Высвобождаемые эндотелием вазокон-
стрикторы (эндотелии) и вазодилататоры (NO) играют важную роль в регуляции почечного крово-тока при повреждении эндотелия.
^ Г. Нервная регуляция. Симпатические волокна, отходящие от спинного мозга на уровне сегментов T/, - L1, достигают почек через чревное и почечное нервные сплетения. Симпатические нервы иннервируют юкстагломерулярный аппарат (P1) и сосуды почек (Q1). Снижение почечного кровотока при стрессе вероятнее всего обусловлено симпатической иннервацией. Стимуляция агадренорецеп-торов повышает реабсорбцию натрия в прокси-мальных канальцах, а а2-адренорецепторов — приводит к снижению реабсорбции натрия и усилению экскреции воды. Действие дофамина осуществляется через специфические почечные D ,-рецепторы. Активация пресинаптических О2-рецепторов на постганглионарных нейронах ингибирует высвобождение норадреналина.
^ Распределение почечного кровотока
Приблизительно 80 % почечного кровотока получают корковые нефроны и лишь 10-15 % — юкста-медуллярные. При определенных условиях происходит перераспределение почечного кровотока от корковых нефронов с короткой петлей Генле в пользу юкстамедуллярных нефронов с длинной петлей Генле. Симпатическая стимуляция, увеличение концентрации катехоламинов и ангиотензи-на II в крови, сердечная недостаточность вызывают перераспределение почечного кровотока из коркового вещества в мозговое. Хотя значение этого явления недостаточно ясно, клинически оно сочетается с задержкой натрия.
^ Влияние анестезии на почечную функцию
Влияние анестетиков на функцию почек в клинических условиях исследовать сложно, потому что трудно отдифференцировать прямое действие анестетиков от опосредованного, а также учитывать значимость таких важных параметров, как характер операции, тип инфузионных растворов, исходную сердечную и почечную функции. Тем не менее существуют некоторые закономерности:
1. Общая и регионарная анестезия вызывают обратимое снижение почечного кровотока, СКФ, диуреза и экскреции натрия.
2. Вышеперечисленные изменения выражены слабее при регионарной анестезии.
3. Большинство изменений опосредовано вегетативными и гормональными влияниями.
4. Эти изменения могут быть частично нивелированы поддержанием адекватного ОЦК и АД.
5. Специфическое нефротоксическое действие оказывают только некоторые анестетики (метоксифлюран и, теоретически, энфлюран и севофлюран) в высоких дозах.
^ Опосредованное влияние анестезии на почечную функцию
Сердечно-сосудистая система
Большинство ингаляционных и неингаляционных анестетиков вызывают депрессию миокарда или вазодилатацию и, соответственно, снижают АД. Приводя к симпатической блокаде, спинномозговая и эпидуральная анестезии способствуют развитию артериальной и венозной вазодилатации, что сопряжено с риском возникновения артериальной гипотонии. Падение АД ниже пределов ауторегу-ляции может вызвать уменьшение почечного кровотока, СКФ, диуреза и экскреции натрия. Инфузия жидкости устраняет артериальную гипотонию, что приводит к нормализации почечной функции.