
23. Пути и закономерности передвижения органических веществ по растению.
В растениях перемещается множество органических соединений, в том числе углеводы, азотсодержащие вещества, регуляторы роста, витамины, вирусы и различные биоциды - гербициды, фунгициды и инсектициды. Большая часть органических соединений перемещается по флоэме, как показывают опыты с кольцеванием, анализы флоэмного сока и эксперименты с радиоактивными метками. Растущее растение можно рассматривать как интегральную систему потребителей питательных веществ, конкурирующих между собой в отношении направления передвижения и количества углеводов и азотсодержащих веществ, двигающихся к различным тканям в зависимости от нужд потребителей и значения разных источников снабжения. Таким образом, любое обсуждение вопроса о передвижении веществ требует количественной характеристики происходящего роста и места, где он наблюдается.
Количество передвигающихся углеводов превышает общее количество всех других растворенных веществ. Углеводы передвигаются преимущественно вниз, но отмечается и значительное движение вверх. Их источниками служат фонды запасных углеводов и продукты текущего фотосинтеза. Количество передвигающихся веществ зависит от потребности в углеводах и близости растущей ткани к источникам снабжения. У большинства древесных растений углеводы передвигаются в виде сахарозы. У немногих семейств важными компонентами флоэмного сока являются рафиноза, стахиоза, вербаскоза. Сахароспирт сорбит обнаружен у яблони и вишни, маннит - у ясеня. Редуцирующие сахара во флоэмном соке не встречаются.
У молодых голосеменных растений умеренной зоны до распускания почек главным потребителем углеводов являются обычно корни. После распускания почек происходит обратное передвижение, когда большее количество углеводов движется к развивающимся побегам. В это время рост камбия является второстепенным потребителем притекающих углеводов. После окончания сезонного роста побегов основными потребителями углеводов являются растущий камбий и корни, возобновляющие рост. Этот обобщенный сезонный ход событий может сильно изменяться в год обильного плодоношения, когда репродуктивные ткани монополизируют основную массу имеющихся углеводов. При одновременном росте корней и побегов соотношение остающихся в побегах и передвигающихся в корни ассимилятов зависит от относительного роста
каждого из этих потребителей и их близости к источникам углеводов.
Растущие плоды часто видоизменяют характер оттока углеводов из листьев разнолистных побегов. Плоды - мощные потребители углеводов, и они часто монополизируют имеющиеся углеводы до такой степени, что вместо нисходящего передвижения наблюдается двустороннее. При этом часть продуктов фотосинтеза направляется вверх, в плоды из листьев, находящихся ниже, а часть передвигается вниз от листьев, расположенных на ветви выше.
У многих взрослых деревьев нижние, угнетенные, ветви не участвуют в снабжении углеводами роста главного ствола. Такие "отрицательные" ветви могут даже не давать достаточно углеводов для образования ксилемы по всей длине самой ветви. Например, у взрослых деревьев дугласии в основании нижних ветвей часто было на 9—10 ксилемных колец меньше, чем в главном стволе у места отхождения этой ветви. Ветви сосны ладанной в нижней половине кроны, имеющие менее трех веточек, или ветви нижней четверти кроны менее чем с пятью веточками совсем не дают углеводов для роста главного ствола.
Регуляторы роста всех классов, за исключением этилена, обнаружены как в ксилеме, так и во флоэме. Передвигаются они, по-видимому, на большие расстояния. Меченые регуляторы роста передвигались из листьев с такой же скоростью, что и другие органические соединения, а вещества, стимулирующие цветение, быстро переходят из листьев в кончики стеблей. Ауксин, гиббереллин, абсцизовая кислота и цитокинины передвигаются из корней по ксилеме. Таким образом, нет сомнений в наличии транспорта регуляторов роста на большие расстояния. Однако физиологическое значение такого дальнего транспорта неясно, так как регуляторы роста могут образовываться во всех органах растений. Поэтому представляется возможным, что синтезом in situ объясняется большинство результатов, приписываемых транспорту из корней и других отдаленных тканей. Для оценки значения транспорта на большие расстояния необходимо дальнейшее изучение источников и потребителей.
Вирусные инфекции возможны только при их прямом введении в определенные ткани, обычно при участии насекомого-переносчика. Все вирусы могут проникать в паренхимные клетки. Некоторые вирусы, по-видимому, адаптированы к определенным тканям, а другие (например, вирусы мозаики) могут инфицировать большинство живых клеток растения. У основной части более специализированных вирусов инфицирование зависит от флоэмной ткани, хотя некоторые, видимо, передвигаются по ксилеме.
Возбудитель вирусной инфекции перемещается по флоэме с током ассимилятов. Поскольку флоэма пронизывает все растение, а транспорт происходит в соответствии с характером изменений потребителей, вирусная инфекция может легко распространиться по всему растению. Инфекция, вызванная вирусами, передвигающимися только по флоэме, не может распространяться за окольцованную часть ствола. В противоположность этому инфекция, вызванная вирусами мозаики, может обойти кольцо через клетки паренхимы в ксилеме окольцованной части ствола. Поскольку инфекционный возбудитель вирусной болезни, передвигаясь по растению, может оставить некоторые его части неинфицированными, вполне вероятно, что возбудитель переносится пассивно. Некоторые вирусные инфекции причиняют серьезный вред флоэме, другие нет.
31. Минеральное питание дерева в лесу.
Минеральное питание имеет большое значение для физиологии дерева, поскольку для нормального роста необходимо достаточное снабжение определенными минеральными элементами. Растениям требуются кислород, вода, двуокись углерода, азот и более 10 минеральных элементов в качестве реагентов или "сырья" для различных синтетических процессов и для
других целей. Люди, не знакомые с физиологией растений, часто называют минеральные питательные вещества "пищей" растений, а в лесоводстве используют термин "подкормка", когда вносят удобрения. Физиологи растений не одобряют применения термина "пища" в этих случаях, так как они обычно ограничивают его значение углеводами, жирами и белками, которые могут быть непосредственно использованы растением в качестве источника энергии, окисляясь при дыхании, или как строительный материал при образовании новых тканей. Минеральные вещества и двуокись углерода обычно рассматриваются физиологами растений скорее как сырье, реагенты или вспомогательные вещества, необходимые для синтеза питательных веществ, чем в качестве самой пищи.
У минеральных питательных веществ в растениях много важных функций. Они могут играть роль структурных компонентов растительных тканей, катализаторов различных реакций, регуляторов осмотического давления, компонентов буферных систем и регуляторов проницаемости мембран. Примерами значения минеральных веществ как составных частей растительных тканей могут быть кальций в клеточных стенках, магний в молекулах хлорофилла, сера в определенных белках, фосфор в фосфолипидах и нуклеопротеидах. Азот, хотя и не относится к минеральным элементам, часто включают в их число, поэтому следует еще раз отметить его значение как компонента белков. Некоторые элементы, в том числе железо, медь и цинк, требуются в очень небольших количествах, но они необходимы, поскольку входят в состав простетических групп или коферментов определенных ферментных систем. Другие элементы, такие, как марганец и магний, функционируют в качестве активаторов или ингибиторов ферментных систем. Некоторые элементы, например бор, медь и цинк, необходимые для функционирования ферментов в незначительных количествах, в более высоких концентрациях очень ядовиты. Токсичность этих и других ионов, таких, как серебро и ртуть, вероятно, связана с их повреждающим действием на ферментные системы.
Значение достаточного снабжения растений минеральным питанием для хорошего роста уже давно оценено в полеводстве и садоводстве, но до последнего времени им пренебрегали лесоводы. Даже сейчас большинство исследований проводится на сеянцах в питомниках, но растущая стоимость земли и более высокие цены на лесоматериалы меняют это положение. Использование новых программ улучшения деревьев способствует получению саженцев с более высокой способностью к росту, поэтому для реализации ростового потенциала саженцев часто оказывается необходимым адекватное снабжение минеральными питательными веществами. Более того, укороченные обороты и использование помимо ствола также и других частей деревьев резко увеличивают скорость извлечения минеральных веществ из почвы. По этим причинам удобрение искусственных насаждений лесных пород может приобрести такое же значение, какое имеет в настоящее время удобрение сельскохозяйственных культур. Дальнейшее изучение специфических потребностей деревьев в минеральных веществах необходимо в качестве основы для практического лесоводства и обеспечения лучшей адаптации деревьев к условиям местообитания.
В растениях найдено более половины элементов периодической системы, и вполне вероятно, что корнями поглощается любой элемент, находящийся в окружающей их среде. По крайней мере 27 элементов были идентифицированы в некоторых образцах древесины сосны веймутовой, а другие безусловно присутствуют в небольших количествах. Не все найденные в деревьях элементы необходимы. В растениях находятся значительные количества алюминия, кремния и натрия, но ни один из этих элементов не считается необходимым.
Минеральный элемент считается необходимым, если, во-первых, без него растения не могут завершить жизненный цикл, и, во-вторых, если он входит в состав молекулы какого-либо необходимого компонента растений.
Недостаток минеральных веществ выз
ывает изменения биохимических и физиологических процессов, в результате чего часто наблюдаются морфологические изменения, или видимые симптомы. Иногда вследствие дефицита рост подавляется до появления других симптомов.
Наиболее существенный результат недостатка минеральных веществ - снижение роста. Однако наиболее заметный эффект - это пожелтение листьев, вызванное уменьшением биосинтеза хлорофилла. Листья, по-видимому, особенно чувствительны к дефициту. При недостатке минеральных веществ у них уменьшаются размеры, изменяется форма или структура, бледнеет окраска, а иногда даже образуются мертвые участки на кончиках, краях или между главными жилками. В некоторых случаях листья собираются в пучки или розетки, а сосновые иглы иногда не могут разделиться и образуется "слившаяся хвоя". Общий признак определенного типа недостаточности минеральных веществ в травянистых растениях - подавление роста стебля и снижение роста листовых пластинок, что приводит к образованию розеток небольших листьев, часто с сетью хлоротичных участков. Видимые симптомы дефицита различных элементов настолько характерны, что опытные наблюдатели могут идентифицировать дефицит по внешнему виду листьев.
Иногда при недостатке минеральных веществ деревья образуют избыточные количества камеди. Это явление получило название гоммоза. Выделение смолы вокруг почек характерно для страдающих от недостатка цинка деревьев сосны замечательной в Австралии. Камедь обнаруживается также на коре плодовых деревьев, страдающих от суховершинности, вызванной недостатком меди. Значительный дефицит часто вызывает гибель листьев, побегов и других частей, т.е. развиваются симптомы, описанные как суховершинность. Отмирание побегов, вызванное недостатком меди, наблюдалось у многих лесных и плодовых деревьев. При отмирании верхушечных побегов яблони, страдающие от дефицита меди, приобретают кустообразный, чахлый вид. Недостаток бора вызывает засыхание верхушечных точек роста и в конце концов гибель камбия у цитрусовых и у сосен, отмирание флоэмы и физиологический распад плодов у других видов. Недостаток одного элемента иногда способствует появлению нескольких различных симптомов, например дефицит бора у яблонь вызывает деформацию и хрупкость листьев, некроз флоэмы, повреждения коры и плодов.
Наиболее распространенный симптом, наблюдаемый при недостатке самых различных элементов, - хлороз, возникающий в результате нарушения биосинтеза хлорофилла. Характер, степень и тяжесть хлороза у молодых и старых листьев зависят от вида растения, от элемента и степени дефицита. Чаще всего хлороз бывает связан с недостатком азота, но он может быть вызван и дефицитом железа, марганца, магния, калия и других элементов. Более того, хлороз может быть вызван не только дефицитом минеральных веществ, но и множеством других неблагоприятных факторов окружающей среды, в том числе избытком или недостатком воды, неблагоприятными температурами, ядовитыми веществами (например, двуокисью серы) и избытком минеральных веществ. Причиной хлороза могут быть и генетические факторы, вызывающие появление различно окрашенных растений: от альбиносов, полностью лишенных хлорофилла, до зеленоватых проростков или проростков сразличной полосатостью и пятнистостью листьев.
В лесных почвах широко распространен недостаток азота, а в некоторых районах наблюдается значительный дефицит фосфора и калия. Эти элементы являются наиболее важными и для минерального питания плодовых деревьев. Большинство
работ по применению удобрений в лесах выполнено на макроэлементах: азоте, фосфоре и калии. Однако у плодовых и декоративных деревьев часто наблюдается дефицит микроэлементов (таких, как железо, цинк, медь и бор), особенно на известковых или песчаных почвах. Дефицит микроэлементов начинает обнаруживаться в лесных насаждениях во всем мире. Без сомнения, будут появляться новые примеры недостаточности микроэлементов по мере того, как экзотические виды станут выращиваться в климатических условиях и на почвах, к которым они мало приспособлены. Опыты с цитрусовыми показывают, что наилучшим способом ликвидации дефицита марганца, цинка и молибдена является опрмскивание листвы. Однако цитрусовые деревья не стоит опрыскивать соединениями железа. В известковые почвы железо следует вносить в виде хелатов.
48. Движение растений. Положительные и отрицательные тропизмы. Настии.
Движения у растений делят на два основных типа: 1) пассивные и 2) активные. Пассивные, или гигроскопические, движения связаны с изменением содержания воды в коллоидах, составляющих оболочку клетки. У цветковых растений гигроскопические движения играют большую роль при распространении семян и плодов. У растущей в пустыне Аравии иерихонской розы в сухом воздухе веточки свёрнуты, а в сыром развёртываются, отрываются от субстрата и переносятся ветром. Плоды ковыля и журавельника благодаря гигроскопичности зарываются в землю. У жёлтой акации зрелый боб высыхает, две его створки спирально скручиваются, а семена с силой разбрасываются. В основе активных движений лежат явления раздражимости и сократимости белков цитоплазмы растений, а также ростовые процессы. Воспринимая влияния окружающей среды, растения реагируют на них усилением интенсивности обмена, ускорением движения цитоплазмы, а также ростовыми и др. движения. Воспринятое растением раздражение передаётся по цитоплазматическим тяжам — плазмодесмам, а затем уже происходит ответ растения как целого на раздражение. Слабое раздражение вызывает усиление, сильное — угнетение физиологических процессов в растении. Активные движения бывают медленные (ростовые) и быстрые (сократительные). К ростовым движениям относятся: тропизмы (раздражение действует в одном направлении и происходит односторонний рост, в результате чего возникает изгиб органа — геотропизм, фототропизм, хемотропизм и др.) и настии (ответ растения на действие раздражителей, не имеющих определённого направления — термонастии, фотонастии и т.д.).
Сократительные движения часто называют тургорными. Эти движения у растений — результат взаимодействия аденозинтрифосфорной кислоты (АТФ) с сократительными белками. Таким образом, механизм сократительных движений растений почти тот же, что и при сокращении мышц человека. К активным сократительным движениям относятся перемещения в пространстве некоторых низших организмов — таксисы, вызываемые, как и тропизмы, односторонним раздражением. К таксисам способны снабженные жгутиками бактерии, некоторые водоросли, антерозоиды мхов и папоротников. Многие водоросли (хламидомонады) обнаруживают положительный фототаксис, антерозоиды мхов собираются в капилляры,
содержащие слабый раствор сахарозы, а папоротников — раствор яблочной кислоты (хемотаксис). К сократительным движениям, связанным, вероятно, с сокращениями белкового вещества цитоплазмы, относятся и сейсмонастии. Близко к сейсмонастиям стоят автономные движения. Так, у семафорного растения Desmodium gyrans сложный лист состоит из большой пластинки и двух меньших боковых пластинок, которые то опускаются, то поднимаются, как семафор; при неблагоприятных условиях (темнота) эти движения прекращаются. У биофитума (Biophytum sensitivum) при сильном раздражении листочки складываются, как у мимозы, совершая ряд ритмических сокращений. При этом, по-видимому, происходит распад АТФ и быстрое её восстановление, что и вызывает непрерывные движения листьев под влиянием раздражителей. Листочки кислицы складываются под влиянием сильного света, темноты, повышенной температуры. К вечеру листочки кислицы складываются, а уже ночью происходит их раскрывание, видимо, после того, как восстановится связь АТФ с сократительными белками. У растений, способных к никтинастическим (Acacia dealbata), сейсмонастическим (Mimosa pudica), а также к автономным Д. (Desmodium gyrans), имеется высокая активность АТФ. У растений, не способных к движениям, она незначительна (Desmodium canadensis). Наибольшим содержанием АТФ отличаются те ткани растений, которые связаны с движениями. Раньше господствовало мнение, что движение листьев мимозы связано с потерей тургора и выходом воды в межклетники в сочленениях листа. В. А. Энгельгардт (1957) предполагает участие АТФ в осмотических явлениях, связанных с Д. листьев мимозы, и дегидратацией её клеток в сочленениях.
Тропизмы (от греч. trоpos — поворот, направление), движения органов растений в ответ на одностороннее действие света, силы тяжести и др. факторов внешней среды в результате более быстрого роста клеток на одной стороне побега, корня или листа. В основе тропизмов (как и настий) лежит явление раздражимости. Начинается процесс с восприятия растением внешнего раздражения, которое индуцирует физиологические различия между двумя сторонами растительного органа, затем следует фаза передачи сигнала и в завершение наступает реакция — изгиб вследствие неравномерности скорости роста двух сторон органа. Наибольшее распространение получила гормональная теория тропизмов, лучше всего демонстрируемая на примере фототропизма и геотропизма. Проросток овса, освещенный с одной стороны, изгибается в сторону источника света вследствие того, что его освещенная сторона растет медленнее, а затенённая быстрее. При этом содержание ауксина оказывается больше в затенённой, быстрее растущей половинке, то есть фототропический изгиб — результат неравномерного распределения ауксина. В горизонтально лежащем стебле ауксин скапливается в нижней части, что приводит к усилению роста этой части стебля и его изгибу вверх (положительный геотропизм); в горизонтально расположенном корне ауксин концентрируется также в нижней части, но при его избытке рост клеток корня, чувствителен к ауксину, затормаживается, в результате чего корень изгибается вниз (отрицательный геотропизм).
Настии (от греч. nastós — уплотнённый) - движения листьев, лепестков и др. органов растений в ответ на воздействие раздражителя (света, температуры или др. фактора), не имеющего определённого направления. Настии — результат неравномерного роста клеток, расположенных на верхней и нижней сторонах органа, или неодинакового изменения тургора в этих клетках. Более интенсивный рост верхней стороны органа вызывает эпинастии (например, при раскрывании почек и цветочных бутонов), нижней стороны — гипонастии (например, при смыкании цветочных покровов по окончании цветения). В зависимости от действующего фактора различают: фотонастии — движения листьев и лепестков, вызываемые изменением интенсивности освещения (наблюдаются у подснежника, кувшинки, одуванчика и др. растений); термонастии — движения лепестков и др. органов, обусловленные изменением температуры (например, у шафрана, тюльпана); никтинастии — движения листьев и лепестков у многих растений, обусловленные сменой дня и ночи; осуществляются в результате сочетания фото- и термонастических движений; сейсмонастии — движения органов растений в ответ на механическое раздражение или сотрясение (движения рылец цветка и тычиночных
нитей у некоторых насекомоопыляемых растений, листьев — у насекомоядных растений, у мимозы стыдливой и др.); хемонастии — движения органов растений, например листьев,— у душистого горошка, томатов и др. растений, обусловленные действием некоторых газообразных веществ (например, незначительных концентраций этилена, ацетилена и CO). Биологическое значение Настий разнообразно: у многих растений они связаны с приспособлением к перекрёстному опылению насекомыми, с защитой цветков от неблагоприятных условий, у насекомоядных — с ловлей насекомых и т.д.
62. На примере представителей порядков отдела зеленые водоросли описать усложнение в процессе эволюции.
Зеленые водоросли — самый многочисленный из всех отделов водорослей. Он насчитывает от 13 до 20 тыс. видов и включает одноклеточные, колониальные и многоклеточные организмы. Все они отличаются чистым зеленым цветом своих слоевищ, сходным с окраской высших растений и обусловленным преобладаем хлорофиллов над другими пигментами.
Вследствие разнообразия строения зеленые водоросли делят на пять классов, в том числе вольвоксовые (основная форма вегетативного тела — подвижные жгутиковые клетки и колонии таких клеток), протококковые (основная форма вегетативного тела — неподвижные клетки с плотной оболочкой и колонии таких клеток), улотриксовые (многоклеточные нитчатые и пластинчатые формы). К зеленым водорослям примыкают наиболее сложно устроенные харовые водоросли, произошедшие от зеленых и выделяемые ныне в самостоятельный отдел.
Вследствие широкого распространения и способности к быстрому размножению вольвоксовые играют существенную роль в круговороте веществ в природе. Они активно участвуют в очистке загрязненных и стоячих вод, в формировании озерного ила — сапропеля, служат источником пищи для многих видов мелких беспозвоночных животных.
Произошли вольвоксовые от примитивных амебоидных форм. Большое значение имеют два направления эволюции вольвоксовых. Первый — усложнение строения слоевища путем дифференциации клеток на вегетативные и репродуктивные. Это один из возможных путей возникновения многоклеточных организмов.
Второй — утрата подвижности в вегетативной фазе жизненного цикла. Эта линия эволюции привела к формированию протококковых водорослей, а от них — через нитчатые и пластинчатые многоклеточные водоросли — к наземным зеленым растениям.
Эволюция зеленых водорослей в водной среде не привела к образованию сложно устроенных и крупных слоевищ с дифференциацией их на ткани. Это произошло при освоении зелеными водорослями суши и развитии высших наземных
растений.
73. Характеристика сфагновых мхов, их роль в образовании торфа.
Сфагнум (Sphagnum), род сфагновых, или торфяных (белых), мхов. Включает 320 видов; в России 42 вида. Преимущественно болотные мхи, произрастающие густыми плотными скоплениями, образующими крупные подушки или сплошные ковры на сфагновых болотах; реже сфагнум встречаются во влажных лесах. Прямостоячий (высотой 10—20 см) мягкий стебель с пучковидно расположенными ветвями и однослойные листья С. содержат большое количество мёртвых водоносных (гиалиновых) клеток с порами, легко впитывающих воду, что обусловливает высокую влагоёмкость сфагнума и способствует быстрому развитию верховых болот в местах, где появляются эти мхи. Стебли сфагнума ежегодно в нижней части отмирают (рост стебля продолжают верхушечные ветви), образуя торф. Распространены преимущественно в тундровой и лесной зонах Северного полушария; в Южном полушарии встречаются высоко в горах, реже на равнинах умеренного пояса.
Таким образом, большую роль играют сфагновые мхи в формировании и жизни болот. Из отмерших остатков сфагнума образуются залежи торфа. Сфагнум растет верхушкой стебля, а его нижняя часть отмирает ("отторфовывается"). Так в течение многих лет образуются огромные залежи торфа. Процесс торфообразования происходит благодаря застойному переувлажнению, отсутствию кислорода и созданию мхами кислой среды. Эти условия оказываются неблагоприятными для процессов гниения, что препятствует разложению сфагнума. Торф - ценное сырье для получения воска, парафина, аммиака, спирта и т.д. Он используется в медицине, в строительной промышленности, в качестве топлива, удобрений.
Значение болот однако не сводится только к образованию в них торфа. На болотах формируется своеобразное сообщество, в котором значительное место занимают клюква, лекарственные растения, обитают животные, в том числе и промысловые. Но главное, болото - это резервуар пресной воды, он, как губка, впитывает в себя все осадки, которые затем вытекают из него в виде ручейков и впадают в реки. Поэтому не всегда можно осушать небольшие болота, так как это может вызвать обмеление и заиление рек, наступление засухи.
Многие мхи угнетают рост и жизнедеятельность таких микроорганизмов, как кишечная палочка, холерный вибрион, золотистый стафилококк, сальмонелла. Эта способность мхов угнетать, подавлять жизнедеятельность других организмов является с
редством их самозащиты, приспособлением к выживанию в условиях острой борьбы за существование.
85. Голосеменные:строениепыльцы, зародышевогомешка, оплодотворение.
Голосеменные - наиболее древняя группа семенных растений. Появились они на Земле свыше 350 млн. лет назад, задолго до возникновения покрытосеменных. Ученые считают, что голосеменные произошли от древних разноспоровых семенных папоротников, которые не дожили до наших дней. Отпечатки семенных папоротников находят в глубоких слоях земной коры.
Все голосеменные растения - деревья или кустарники. Нередко они достигают огромных размеров, как некоторые хвойные (секвойя, Мамонтове дерево высотой свыше 110 м). В отличие от покрытосеменных голосеменные не имеют цветка и плода. Семена у них образуются в шишках и располагаются на поверхности чешуек, за что растения и получили свое название - голосеменные.
Отдел голосеменных объединяет 6 классов: семенные папоротники, саговниковые, беннетитовые, гнетовые, гинкговые, хвойные. К настоящему времени семенные папоротники, беннетитовые полностью вымерли и об их существовании свидетельствуют лишь отпечатки и окаменелости различных органов этих растений. Из гинкговых до наших дней дожил всего один вид - гинкго двулопастный.
Самый многочисленный и наиболее распространенный класс современных голосеменных - хвойные. Их насчитывают всего около 560 видов, несмотря на то, что по древности хвойные превосходят все ныне живущие голосеменные. Самые древние из хвойных - растения семейства сосновых (сосна, ель, пихта и др.). Они существуют более 300 млн. лет и сохранили основные черты строения своих предков.
Особенности строения и жизнедеятельности голосеменных можно рассмотреть на примере сосны обыкновенной. Это дерево высотой до 40 - 50 м, живет до 400 лет, растет повсюду: на бедных песчаных почвах, сфагновых болотах, известняковых склонах.
Размножение хвойных рассмотрим на примере сосны (рис. 1).
Сосна — обоеполое (однодомное) растение. Весной у основания ее молодых побегов образуются пучки мужских шишек. Каждая шишка состоит из оси, одетой многочисленными чешуйками. На нижней стороне чешуек располагаются два
микроспорангия (пыльцевых мешка). В них в результате митотических делений возникают гаплоидные микроспоры, где развивается сильно редуцированный гаметофит, состоящий из нескольких клеток. В результате последовательных делений формируется пыльцевое зерно. Оно состоит из оболочек, из вегетативной клетки, образующей пыльцевую трубку, и генеративной клетки (из нее формируется спермий).
Женские шишки также состоят из оси, на которой расположены семенные чешуйки. Женские шишки возникают на концах побегов текущего года. Чешуи состоят из двух частей: наружной — кроющей и внутренней —: семенной. На семенных чешуях формируются по два семязачатка. Семязачаток включает мегаспорангий, который одет оболочкой и имеет канал для проникновения пыльцевой трубки. В мегаспорангий возникает крупная мегаспора, которая путем ряда клеточных делений превращается в женский гаметофит. В гаметофите образуется яйцеклетка, а сам он превращается в гаплоидный эндосперм.
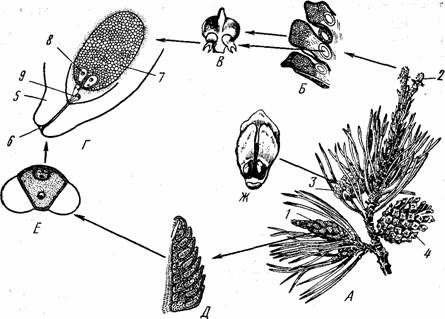
Рис. 1. Схема опыления у сосны: А — ветка с шишками; Б — женская шишка в разрезе; В — семенная чешуя с семязачатками; Г — семязачаток в разрезе; Д — мужская шишка в разрезе; Е — пыльца; Ж — семенная чешуя с семенами:
1 — мужская шишка, 2 — молодая женская шишка, 3 — шишка с семенами, 4 — шишка после высыпания семян, 5 — покров, 6 — семявход, 7 — эндосперм, 8 — яйцеклетка, 9 — пыльцевая трубка со спермиями
Опыление у сосны происходит в конце весны или начале лета. Семенные чешуйки женской шишки раздвигаются. Пыльца, разносимая ветром, попадает в щели между семенными чешуйками. Через канал мегаспорангия выделяется так называемая «опылительная жидкость», выступающая на верхушке семязачатка в виде капли. Пыльцевые зерна погружаются в нее, после чего семенные чешуи сближаются и остаются сомкнутыми до созревания семян. Оплодотворение у сосны происходит только на слудующий год — через 12—14 мес. после опыления. Пыльца прорастает, пыльцевая трубка проникает через канал мегаспорангия и достигает яйцеклетки. Один из двух спермиев сливается с яйцеклеткой, другой погибает. Из зиготы развивается зародыш. Из покрова семязачатка образуется кожура семени. После созревания семян чешуйки шишки
расходятся и семена высыпаются.
99. Опишите семейства со сростнолепестным околоцветником, укажите представителей.
Сростнолепестные, подкласс покрытосеменных двудольных растений; то же, что спайнолепестные. Спайнолепестные, сростнолепестные, подкласс покрытосеменных двудольных растений, объединяющий семейства, для представителей которых характерно срастание лепестков. Срастание может быть частичным — касаться лишь оснований лепестков, либо более или менее полным — доходить до их середины и выше вплоть до срастания по всей длине. У большинства сростнолепестных сросшаяся базальная часть лепестков образует трубку, а верхние части имеют вид лопастей, долей, сегментов или зубчиков. Для сростнолепестных характерны также: сростнолистная чашечка; циклический цветок; срастание на более или менее значительном протяжении тычиночных нитей с цветочной трубкой; 1 (а не 2, как у большинства раздельнолепестных) покров семязачатка. Выделение сростнолепестных (Sympetalae) в противоположность раздельнолепестным было предложено в 1864 нем. ботаником А. Брауном, а в 1892 нем. ботаник А. Энглер предложил для сростнолепестных термин «вторичнопокровные» (Metachlamydeae), считая их более высокоорганизованными, чем раздельнолепестные и первичнопокровные. В большинстве современных филогенетических систем сростнолепестных в особый подкласс двудольных не выделяют. Охарактеризуем некоторых представителей сростнолепестных.
Мареновые (Rubiaceae), семейство двудольных растений. Деревья, кустарники, полукустарники, лианы и травы. Листья цельнокрайные или иногда зубчатые, супротивные, часто мутовчатые, с прилистниками (нередко листовидными). Цветки большей частью обоеполые, 4—5-членные, правильные, в цимозных соцветиях, реже одиночные. Чашечка сростнолистная, обычно слабо развита; венчик сростнолепестный. Тычинки по числу лопастей венчика. Плоды — коробочки или распадающиеся двусемянки, реже ягодовидные или костянковидные. Около 500 родов (6500 видов), встречающихся повсеместно, но главным образом в тропиках и субтропиках. В России 13 родов (около 200 видов). К мареновым относятся кофейное дерево, лекарственные (хинное дерево, ипекакуана и другие), красильные (марена, моринда), декоративные (гардения, бувардия), дубильные и другие хозяйственно-ценные растения.
Ластовневые, ласточниковые (Asclepiadaceae), семейство двудольных растений. Лазящие, вьющиеся или прямостоячие кустарники или полукустарники и травы, изредка деревья. Листья большей частью супротивные, цельные и без прилистников. Цветки обоеполые, обычно правильные, 5-членные. Венчик сростнолепестный, часто с так называемой коронкой. Тычинки с гинецеем образуют гиностегий. Пыльцевые зёрна в поллиниях, реже соединены по 4 в тетрады. Своеобразное строение цветка приспособлено к перекрёстному опылению насекомыми. Плод из 2 листовок, несколько раздвинутых и напоминающих хвост ласточки (отсюда название); семена обычно с хохолком из шелковистых волосков. Для ласточниковых характерно наличие млечного сока (часто ядовитого) и интраксилярной флоэмы (внутреннего луба). Около 290 родов (2000 видов), главным образом в тропических и отчасти в субтропических и умеренных областях. В России 8—9 родов (около 40 видов). Важнейшие роды: ваточник, ластовень, обвойник и др. Среди ласточниковых много ядовитых, лекарственных и декоративных растений (кондуранго, восковой плющ, стапелня и др.); некоторые ласточниковые дают красящие вещества и волокно (виды Marsdenia, Calotropis и др.).
Вьюнковые (Convolvulaceae), семейство двудольных растений. Травы, полукустарнички, кустарники, редко небольшие деревья; многие — вьющиеся растения. Цветки часто крупные, одиночные или в соцветиях, обычно обоеполые, 5-членные (редко 4-членные). Венчик сростнолепестный, правильный, слегка 5-лопастный или с цельным краем, воронковидный, реже трубчатый или колокольчатый. Завязь верхняя. Плод — коробочка, изредка ореховидный. Свыше 50 родов (1500 видов), главным образом в тропиках и субтропиках обоих полушарий; наибольшее хозяйственное значение имеют батат и ялапа. В России около 40 видов из 4 родов: кресса, ипомея, вьюнок и повой. В садах юга России разводят тропические декоративные растения рода фарбитис.
Список использованной литературы
1. Алексеев Ю.А., Вахромеева М.Г., Денисова Л.В., Никитина СВ. Лесные травянистые растения. М.: Агропромиздат, 1988.
2. Власова Н.П. Практикум по лесным травам. М.: Агропромиздат, 1986.
3. Васильев А.Е., Воронин Н.С., Еленевский А.Г. и др. Ботаника. Морфология и анатомия растений. М.: Просвещение,1998.
4. Грин Н., Стаут У., Тейлор Д. Биология. Т. 1-3. М.: Мир, 1990.
5. Гэлстон А., Девис П., Сэттер Р. Жизнь зеленого растения. М.: Мир, 1983.
6. Жизнь растений. Т.1-6 М.: Просвещение, 1980.
7. Крамер П., Козловский Т. Физиология древесных растений. М.: Лесная промышленность, 1983.
8. Матвеева Г.В., Тарабрин А.Д. Ботаника. М.:Агропромиздат, 1989.
9. Полевой В.В. Физиология растений. М.: Высшая школа, 1989.
10. Родионова А.С, Барчукова М.В. Ботаника. М.:Агропромиздат, 1990.
11. Родионова А.С. Лесная ботаника. М.: Лесная промышленность, 1980.
12. Рейвн П., Эверт Р., Айкхорн С. Современная ботаника. Т.1-2. М.: Мир, 1990.
13. Сибирякова М.Д. Типы леса лесорастительных районов Европейской части СССР с иллюстрацией подлесной флоры. М.: Гослесбумиздат, 1962.
14. Хржановский В.Г., Пономаренко С.Ф. Ботаника. М.: Агропромиздат, 1988.