
В засушливых местообитаниях листья могут служить для запасания воды (алоэ), которую они очень медленно расходуют. У таких растений воды испаряется очень мало благодаря малому числу устьиц и сильному развитию кутикулы. Приспособлениями к существованию в засушливых местах служат также утолщения стенок эпидермиса и образование покровными клетками воскового налета, густое опушение листьев. У некоторых растений устьица находятся в углублениях листа, что также уменьшает испарение. Листья многих злаков в жаркое время дня свертываются в трубку. Устьица оказываются на внутренней поверхности трубки и изолируются от окружающего сухого воздуха. В полости трубки концентрация водяных паров повышается, что приводит к ослаблению транспирации.
В процессе приспособления к условиям окружающей среды листья помимо основных приобретают дополнительные функции. Защитную и влагосберегающую роль играет преобразование листьев в колючки у барбариса и кактуса. У гороха видоизменением листьев являются усики, с помощью которых растение цепляется за опору. У репчатого лука листья превратились в сочные чешуи, запасающие питательные вещества. Тонкие наружные чешуи играют защитную роль.
Обитание растений на почвах, бедных азотистыми веществами, привело к формированию из листьев ловчих аппаратов, служащих для захвата и переваривания насекомых (росянки, некоторые лианы).
23. Простые и сложные листья. Опишите и нарисуйте основные формы сложных листьев, приведите примеры.
Лист (folium), орган высших растений, выполняющий функции фотосинтеза и транспирации, а также обеспечивающий
газообмен с воздушной средой и участвующий в др. важнейших процессах жизнедеятельности растения.
Форма листьев очень разнообразна. Различают лист простой — с черешком и одной пластинкой, и лист сложный — с главным черешком, или главной осью (рахисом), и черешками (осями) последующих порядков, к которым прикрепляются листочки; части, слагающие типичный сложный лист, снабжены сочленениями. Пластинка листа по форме (очертанию) бывает округлой, эллиптической, ланцетовидной, ромбовидной, яйцевидной и т.п. (рис. 2). Лист может быть также игловидным, шиловидным, щетиновидным, цилиндрическим и иной формы. Различают, кроме того, листья чешуевидной, щитовидной, воронковидной и др. форм. Для классификации листьев большое значение имеет форма основания пластинки (сердцевидная, копьевидная, почковидная и др.), характер очертания её верхушки (заострённая, тупая и т.п.) и особенности её края (зубчатый, пильчатый, городчатый, цельный и т.д.). В зависимости от характера и глубины расчленения пластинки различают листья лопастные, раздельные или рассеченные. Большое значение (особенно для исследования ископаемых растений) имеет характер расположения жилок в пластинке, т. е. жилкование (перистое, пальчатое, дланевидное, дуговидное и др.). Лист может быть голым, опушенным, морщинистым, гладким, а также плёнчатым, кожистым или сочным. Средняя длина листа 3—10 см, однако размеры его варьируют от нескольких мм до 20 м (у пальмы рафии смолистой).

Рис. 2. Лист: 1 — игловидный; 2 — линейный (видны узел, влагалище, язычок и пластинка); 3 — продолговатый; 4 — ланцетовидный; 5 — овальный; 6 — эллиптический, дуговидножилковатый, цельнокрайный; 7 — округлый; 8 — яйцевидный, перистожилковатый, зубчатый; 9 — обратнояйцевидный; 10 — ромбовидный; 11 — лопатчатый; 12 — сердцевиднояйцевидный, городчатый; 13 — почковидный; 14 — стреловидный; 15 — копьевидный; 16 — перистолопастный; 17 — пальчатолопастный, пальчатожилковатый; 18 — пальчатораздельный с пильчатыми долями; 19 — пальчаторассечённый; 20 — лировидный; 21 — тройчатосложный; 22 — пальчатосложный; 23 — парноперистосложный с прилистниками и усиками; 24 — непарноперистосложный с прилистниками; 25 — дваждыперистосложный; 26 — многократноперистосложный; 27 — прерывчатоперистый; 28 — чешуйчатый.
31. Сущность вегетативного размножения и возобновления растений. Биологическое и хозяйственное значение вегетативного размножения и возобновления.
Вегетативное размножение - образование нового организма из части материнского. Способы вегетативного размножения, наблюдаемые в природе, разнообразны. Простейший из них — восстановление организма из одной вегетативной клетки в результате её последовательного деления и дифференциации образующихся клеток.
Вегетативное размножение свойственно микроорганизмам, низкоорганизованным животным, почти всем растениям. У животных вегетативное размножение осуществляется путём почкования (например, у губок, кишечнополостных, мшанок) или деления (например, у простейших); у одноклеточных растений (водоросли, грибы и др.) — чаще путём деления, реже почкования, у низших многоклеточных растений распадением тела на отдельные участки, способные к регенерации (восстановлению). Высшие растения могут размножаться корневищами, отпрысками, луковицами, клубнями и др. вегетативными органами. У многих культурных растений вегетативное размножение — единственно возможный путь сохранения ценных сортовых признаков и свойств.
У высших растений существует размножение отводками (пихта, рододендроны, дикие формы винограда и т.д.), то есть
частями органов, связь которых с материнским растением поддерживается до тех пор, пока они не начинают питаться самостоятельно; плетями или усами (земляника, ежевика, батат, лютик ползучий, будра и др.); корневыми отпрысками (многие лиственные и хвойные деревья и травы — рябина, роза, ольха, таусагыз, осот полевой и др.). Луковицами, клубнями размножаются многие овощные и декоративные растения (лук, тюльпан, лилия, картофель, батат и многие др.), корневищами, как правило, — многолетние растения (ландыш, мята, спаржа, бамбук и многие сорные растения). В растениеводстве широко распространено размножение черенками (стеблевыми, корневыми и листовыми). Естественное размножение листьями свойственно бриофиллуму, у которого образовавшиеся на листовых пластинках побеги опадают и укореняются в почве.
В плодоводстве применяют корнесобственные способы вегетативного размножения (размножение отводками и черенкование) и прививку. Отводок — часть наземного стебля с почками, не отделяемая на период укоренения от материнского растения. Стебли, подлежащие укоренению, окучивают. Укоренившиеся стебли отделяют от материнского растения, разрезают на части (имеющие корни) и сразу высаживают на плантации или доращивают в питомнике (1—2 года). Многие виды с.-х. и декоративных растений (ежевика, плющ, сумах, скумпия, рододендрон и др.) размножаются естественными отводками. В плодоводстве чаще всего применяют размножение вертикальными и горизонтальными отводками; так размножают многие сорта крыжовника, некоторые подвои яблони, в декоративном садоводстве — сирень, калину, жимолость и др. Многие сорта ежевики и ежевикообразной малины размножают верхушечными отводками.
При черенковании используются части растения (черенки), способные при определенных условиях восстанавливать корни (на стеблях), или почки (на корнях), или почки и корни (на листьях). Различают черенки стеблевые, листовые и корневые. Стеблевые черенки могут быть без листьев (зимние), с листьями (летние, или зелёные), травянистые и деревянистые. Безлистные черенки обычно заготавливают осенью, чаще из однолетних побегов, которые разрезают на части длиной 20—30 см, высаживают осенью или весной, сохраняя их прикопанными в подвале, в снегу. Зимними черенками размножают виноград, некоторые сорта крыжовника, некоторые типы подвоев яблони и сливы, айву, инжир, гранат, спирею, жасмин, тополь, иву и др. Для зелёных черенков используют облиственные побеги, разрезая их на части длиной 5—10 см, иногда срезают со стебля часть коры с древесиной и одной почкой (листопочковые черенки). Зелёными черенками размножают крыжовник, вишню, сливу, айву, облепиху, маслину, многие декоративные кустарники и травянистые цветочные растения — хризантему, герань, гвоздику, георгину, флокс и др. Процесс корнеобразования у зелёных черенков многих растений стимулируют с помощью ростовых веществ. У вишни, сливы, сирени и др. зелёные черенки легче укореняются в период, когда черенкуемые побеги находятся в фазе интенсивного роста в длину; у яблони, крыжовника и др. — в конце этой фазы, а у чёрной смородины и некоторых сортов крыжовника — в течение всей фазы роста. Большое значение при размножении зелёными черенками имеют температура (оптимальная 20—25°С), влажность воздуха и субстрата, световой режим. При зелёном черенковании применяют искусственный туман, создаваемый автоматизированными системами над местом
укоренения черенков. В атмосфере тумана резко возрастает укореняемость черенков, в 2—3 раза сокращаются затраты труда.
Листовой черенок — это лист или его часть. Лист сажают в среду укоренения (чаще в песок) черешком или укладывают нижней стороной, слегка надрезая выпуклые жилки листа. Затем слегка присыпают песком, поливают. Листовыми черенками размножают бегонию, седум, узамбарскую (африканскую) фиалку и др.
Корневые черенки — части корня; заготавливают их обычно осенью, длиной 10—15 см, хранят прикопанными в песке и торфе, весной высаживают в плодородную почву. Корневыми черенками размножают малину (красную), ежевику, молодые сеянцы яблони, некоторые сорта вишни, сливы и др.
Длительность существования леса на определённой территории зависит, в частности, от степени его возобновления. Различают естественное возобновление (семенное или вегетативное, порослевое) и искусственное — посевом семян или посадкой древесных пород разного состава и возраста. Всходы в лесу представлены видами растений всех ярусов, подрост — теми, которые в благоприятных условиях могут выйти во второй или первый ярусы и т. о. обеспечить смену взрослого древостоя. На всходы и подрост главных древесных пород оказывают влияние деревья верхних ярусов: они вследствие сомкнутости крон отнимают у всходов и подроста значительную часть нужного им света, атмосферной влаги, питательных веществ и таким образом задерживают их развитие, что приводит к сильному изреживанию и даже к полной гибели всходов или подроста. Однако некоторое затенение всходов и подроста имеет и положительное влияние: обеспечивается защита их от весенних и раннелетних заморозков, вымокания, ускоряется рост и вынос кроны в верхние ярусы, очищение от сучьев и пр. На лесных опушках любого участка леса, который контактирует с другими типами растительного покрова, виды древесных пород могут внедряться (чаще всего вегетативным способом) в др. типы растительности. Особенно часто это наблюдается в местах контакта леса с лугом, степью, болотом, а также в долинах рек или на их водоразделах, когда лесные породы могут продвигаться далеко на север (в тундру) или на юг (в степь). Сбрасываемые деревьями хвоя, листва ложатся плотным слоем на почву и вследствие медленного их разложения задерживают появление всходов древесных пород, ослабляют травяной покров. Такое же действие оказывает разрастающийся иногда на почве мохово-лишайниковый покров, особенно в хвойных лесах. На процессы возобновления леса оказывают влияние и животные.
48. Строение семян и всходов многодольных, двудольных и однодольных растений. Рисунки.
Семя растения (semen), орган семенных растений, выполняющий функции их воспроизведения, расселения и переживания
неблагоприятных условий. В ходе эволюции разноспоровых высших растений семя возникло в связи с изменением условий существования (климатических и др.) как фаза в онтогенезе между половым процессом, происходящим внутри мегаспорангия (семязачатка, семяпочки), и взрослым растением. Поэтому семя развивается обычно после оплодотворения (иногда, вторично, без оплодотворения — апомиксис), а число семян, образующихся на одном растении, не может превышать число мегаспорангиев, возникших в результате бесполого размножения. Тип семени определяется типом семяпочки, из которой образуется семя, и её ориентировкой, хотя в некоторых случаях после оплодотворения из-за неравномерного роста молодого семени его тип может отличаться от типа семяпочки. На поверхности семя обычно можно различить семенной рубчик — место отделения зрелого семени от семяножки, след микропиле на корневом конце в виде ямки, след халазы на семенодольном конце в виде отличающегося по окраске пятна, бугорка или ямки, и семяшов между семенным рубчиком и халазой, возникающий в результате прирастания к семяпочке верхней части семяножки. Нередко семя имеет выросты — ариллус (кровельку, присемекник), ариллоиды, строфиоли, обеспечивающие мирмекохорию. Поверхность семени бывает гладкой и блестящей (горох), сетчато-ямчатой (белена), шиповатой (куколь), бугорчатой (ясколка), чешуйчатой (борец) и т. д. У некоторых семян образуются крылья (эремурус, бигнония), волоски по всей поверхности (хлопчатник).
Семя. состоит из семенной кожуры (спермодермы), зародыша (молодого спорофита) и у многих растений из тканей с запасными питательными веществами, развивающихся вне зародыша, — перисперма или эндосперма (рис. 3).
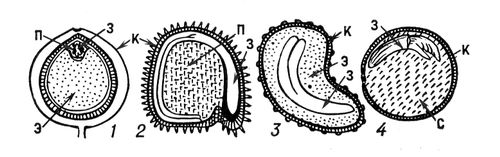
Рис. 3. Строение семян: 1 — чёрного перца (с эндоспермом и периспермом); 2 — куколя (с периспермом); 3 — мака (с эндоспермом); 4 — гороха (без эндосперма и без перисперма); з — зародыш; п — перисперм; э — эндосперм; к — кожура семени; с — семядоля.
Прорастание семени начинается с поступления в него воды: оно набухает, и семенная кожура лопается. Из-под нее появляется кончик зародышевого корешка, который начинает быстро удлиняться, загибаясь под действием силы тяжести вниз. Нижняя, т.е. находящаяся между корешком и семядолями, часть зародышевого стебелька (подсемядольное колено) выходит из-под семенной кожуры и остатков эндосперма «спиной вперед», подтягивая за собой семядоли. Потом стебелек распрямляется и часто выносит семядоли на поверхность, где они расправляются и где начинается фотосинтез. Иногда на семядолях сохраняются в виде колпачка обрывки семенной кожуры. На этой стадии проросток уже способен сам поглощать из почвы воду с минеральными солями и осуществлять фотосинтез, т.е. становится независимым от прочих частей семени и плода. Иногда, впрочем, семядоли остаются под землей, как у гороха, и первой трогается в рост получающая от них пищу почечка. Из почечки развивается надземный стебель и первые фотосинтезирующие листья.
Однодольные, односемядольные, или односеменодольные - класс покрытосеменных растений, характеризующийся наличием у зародыша одной семядоли (отсюда название). У однодольных, в отличие от двудольных, проводящие пучки обычно располагаются беспорядочно, закрытые — лишены образовательной ткани (камбия), вследствие чего стебель и корень в толщину не растут (у немногих однодольных вторичный рост происходит, но, как правило, лишь в результате появления по периферии органа вторичной образовательной ткани); листья обычно с параллельным или дуговидным жилкованием. Корешок зародыша, как правило, отмирает, и вместо главного корня развивается система придаточных корней; стебли разветвлены слабо; листовая пластинка чаще не расчленена, поэтому листья обычно простые, цельные и цельнокрайные. Лишь у некоторых видов однодольных имеется камбий, вскоре прекращающий свою деятельность.
Двудольные, двусемядольные, двусемянодольные - класс покрытосеменных растений, характеризующийся наличием у зародыша двух боковых супротивных семядолей (отсюда название). У двудольных, в отличие от однодольных, проводящие пучки располагаются кольцеобразно, а между древесиной (ксилемой) и лубом (флоэмой) находится особая образовательная ткань — камбий, обеспечивающая вторичное утолщение; листья, как правило, с сетчатым жилкованием: число частей цветка (чашелистиков, тычинок и плодолистиков) обычно кратно 4 или 5. Корешок зародыша чаще всего превращается в главный корень, способный к долголетнему существованию; листовая пластинка часто расчленена, края сё выемчатые или зубчатые. Прорастание семени фасоли представлено на рис. 4.
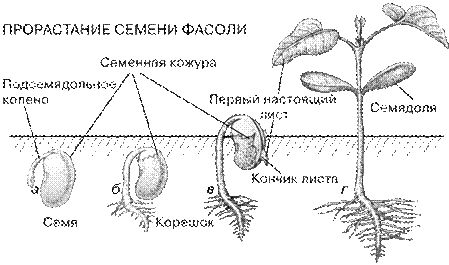
Рис. 4. Прорастание семени фасоли
62. Мейоз. Чем отличается мейоз от митоза? В каких частях растений происходит мейоз, а в каких митоз?
Мейоз (от греч. méiosis — уменьшение), редукционное деление, деления созревания, способ деления клеток, в результате которого происходит уменьшение (редукция) числа хромосом в два раза и одна диплоидная клетка (содержащая два набора хромосом) после двух быстро следующих друг за другом делений даёт начало 4 гаплоидным (содержащим по одному набору хромосом). Восстановление диплоидного числа хромосом происходит в результате оплодотворения. Мейоз — обязательное звено полового процесса и условие формирования половых клеток (гамет). Биологическое значение мейоза заключается в поддержании постоянства кариотипа в ряду поколений организмов данного вида и обеспечении возможности рекомбинации хромосом и генов при половом процессе. Мейоз — один из ключевых механизмов наследственности и наследственной изменчивости. Поведение хромосом при мейозе обеспечивает выполнение основных законов наследственности.
Митоз (от греч. mítos — нить), кариокинез, непрямое деление клетки, наиболее распространённый способ воспроизведения (репродукции) клеток, обеспечивающий тождественное распределение генетического материала между дочерними клетками и преемственность хромосом в ряду клеточных поколений. Биологическое значение митоза определяется сочетанием в нём удвоения хромосом путём продольного расщепления их и равномерного распределения между дочерними клетками. Началу митоз предшествует период подготовки, включающий накопление энергии, синтез дезоксирибонуклеиновой кислоты (ДНК) и репродукцию центриолей. Источником энергии служат богатые энергией, или так называемые макроэргические, соединения.
В зависимости от места митоза в жизненном цикле организмов различают 3 типа. Митоз гаметный, или терминальный митоз (у всех многоклеточных животных и ряда низших растений), происходит в половых органах и приводит к образованию гамет. Зиготный, или начальный, митоз (у многих грибов и водорослей), происходит в зиготе сразу после оплодотворения и приводит к образованию гаплоидного мицелия или таллома, а затем спор и гамет. Споровый, или промежуточный, митоз (у высших растений), имеет место накануне цветения и приводит к образованию гаплоидного гаметофита, в котором позднее образуются гаметы.
73. Основные ткани. Строение, функции и размещение в теле растения запасающих тканей.
Наибольший объем в растении занимают основные ткани. Снаружи они защищены покрывными тканями, а изнутри пронизаны проводящими и механическими пучками. Образующие их клетки отличаются обычно паренхимным строением, из-за этого основные ткани нередко называют основной паренхимой. Основная ткань, в противоположность другим очень богата межклеточными пространствами. Нередко межклетники во много раз превышают размеры самих клеток.
Основные ткани являются преимущественно питающими тканями, хотя по своему существу могут выполнять многие функции, начиная от ассимиляционной, кончая выделительной.
В зависимости от специализации, основную паренхиму делят на ассимиляционную, вентиляционную, всасывающую, запасающую и выделительную.
Ткани, основной функцией которых является работа ассимиляции, то есть фотосинтез, объединяют в систему ассимиляционных тканей.
У высших растений они имеют обычно зеленую окраску. Поэтому могут быть названы зеленой паренхимой или хлорофиллоносной паренхимой (короче - хлоренхимой).
Ассимиляционная ткань устроена достаточно просто и состоит из однородных тонкостенных клеток.
Хлоропласты в клетках хлоренхимы обычно расположены в один ряд в постенном слое цитоплазмы. Центральная часть полости клетки занята крупной вакуолью.
Доступ углекислоты к клеткам хлоренхимы облегчается тем, что в ней имеется развитая система межклетников, сообщающаяся с атмосферой. Наличие межклетников является характернейшей особенностью ассимиляционной ткани. Межклетники обеспечивают газообмен с окружающей средой.
В соответствии с тем, что работа ассимиляции происходит за счет солнечной энергии, хлоренхима располагается в местах, наиболее доступных свету: она находится в надземной части растений непосредственно под кожицей листьев и
стеблей.
Нередко хлоренхима дифференцирована на столбчатую (палисадную) и губчатую ткань. Палисадная ткань обычно состоит из удлиненных клеток цилиндрической формы, расположенных перпендикулярно к поверхности органа. Межклеточники в палисадной ткани развиты слабо. Палисадная ткань содержит большое количество хлоропластов, здесь происходят световые реакции фотосинтеза.
Губчатая ткань построена из округлых или неопределенной формы клеток, образующих рыхлую сложную сетчатую систему. Межклетники хорошо развиты. Здесь протекает газообмен и темновая стадия фотосинтеза.
Практически во всех тканях имеются межклетники, образующие единую систему. Межклетники через проходные отверстия или устьица сообщаются с атмосферой. Однако газовый состав в межклетниках сильно отличается от газового состава атмосферы.
Если в растениях образуется ткань с очень большими межклетниками, нередко превышающими размеры самих клеток, и если вентиляционная функция такой ткани выступает на первое место, то ее называют аэренхимой.
Как правило, в состав аэренхимы входят механические клетки, придающие этой рыхлой ткани дополнительную прочность.
Особенно развита аэренхима у водных и болотных растений, в условиях, где затруднен нормальный газообмен.
Поглощенные растением синтезированные вещества могут откладываться в виде запасов. К накоплению запасных веществ в той или иной мере способны все живые клетки, но в том случае, когда запасающая функция гипертрофированна (выступает на первое место), говорят о запасающих тканях.
Запасы могут храниться длительное время, например, зимние запасы крахмала в корнях и клубнях двулетних и многолетних растений.
В других случаях запасы почти непрерывно потребляются и вновь пополняются в период вегетации.
В строении запасающих клеток встречается большое разнообразие.
Некоторые запасающие ткани приспособлены к накоплению воды. Их называют водоносными. Как правило, клетки водоносных тканей чрезвычайно гигроскопичны (т.е. способны быстро впитывать влагу) и имеют тонкие оболочки.
Наличие водоносных тканей характерно для некоторых ксерофитов - растений, приспособленных переносить длительный засушливый период. Чаще всего водоносные ткани встречаются в листьях. Они являются резервуарами влаги. При подсыхании растения водоносные клетки передают воду главным образом ассимиляционным тканям.
Мощно развитая водоносная ткань встречается у суккулентов - растений с мясистыми сочными листьями и стеблями: агавы, алоэ, кактусы, молочаи.
Развитой водоносной тканью снабжены клубневидные вздутия стеблей многих эпифитных орхидных.
Однако подлинными рекордсменами в запасании влаги являются сфагновые мхи. Они могут накапливать такое количество влаги, которое в 40 - 50 раз превышает их сухой вес. Такая высокая гигроскопичность сфагнов связана с особенностями их анатомического строения. В листьях сфагновых мхов имеются две группы клеток: особые гиалиновые клетки, имеющие большие или многочисленные поры и накапливающие воду и обычные хлорофиллоносные клетки. Гиалиновые клетки занимают большую часть объема. Нередко крупные водоносные клетки имеют спиральные или кольчатые утолщения оболочек, придающих клеткам дополнительную прочность и предохраняющие их от слияния.
Значительно шире распространены ткани, запасающие пластические вещества. Обычно запасы накапливаются в полостях
клеток, реже в их оболочках.
Наиболее распространенные запасные вещества: сахар, инулин, аминокислоты, белки, крахмал.
В клеточных стенках обычно откладываются гемицеллюлозы. В этих случаях стенки чрезвычайно сильно утолщаются. В качестве примеров можно назвать запасающие ткани эндосперма семян кофе и финиковой пальмы.
Таким образом при митозе: у эукариот существует два механизма клеточного деления, связанные с гарантированной передачей полного набора наследственного материала (ДНК). В процессе подготовки к делению каждая хромосома удваивается. Диплоидный набор хромосом в соматических клетках обозначается 2п; количество ДНК обозначается С; таким образом, перед началом деления проходит полная репликация ДНК и клетка содержит 2п, 4с (дочерние нити ДНК). При делении специальный механизм осуществляет попарное сближение гомологичных хромосом, их расхождение к полюсам и деление клетки на дочерние, содержащие по 2с (удвоенные хроматиновые нити). После расхождения дочерние нити ДНК превращаются в хромосомы, образуя комплексы с белками (хроматин) и характерные структуры. В дочерних клетках - снова дигоюидный набор хромосом (2п).
Мейоз же - редукционное деление, при котором после однократной репликации ДНК (2п, 4с) следует два деления клетки (In, 2c, затем In, 2c); в результате дочерние клетки получают полный набор генов, но становятся гаплоидными. Этот механизм клеточного деления в природе встречается только при подготовке к половому размножению, при образовании половых клеток (гамет). При слиянии гамет в процессе оплодотворения получает опять диплоидный набор хромосом.
85. Строение коры ветки лиственной породы. Рисунок поперечного среза.
Кора - периферическая, лежащая кнаружи от камбия часть стеблей и корней растений; слагается из различных по строению и происхождению тканей. Первичная кора формируется в конусе нарастания из наружных слоев меристемы между первичными покровными тканями (эпидермисом в стеблях и эпиблемой в корнях) и центральным цилиндром. Она состоит обычно из паренхимы. Прилегающий к центральному цилиндру слой первичной коры (эндодерма) не всегда ясно выражен. Вторичная кора (вторичная флоэма, или луб) образуется камбием, состоит из ситовидных трубок, сопровождающих клеток, лубяной паренхимы, механических волокон; служит для проведения продуктов фотосинтеза вниз по растению; функционирует как проводящая ткань 1—2 года.
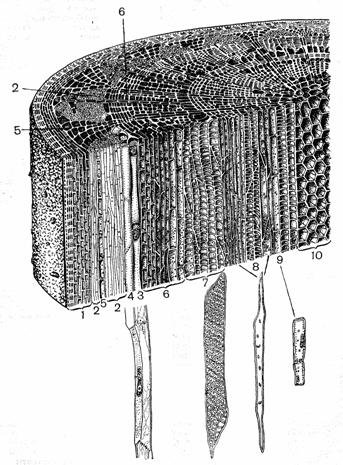
Рис. 5. Клеточное строение однолетнего стебля липы. Продольный и поперечный срезы:
1 — система покровных тканей (снаружи внутрь: один слой эпидермиса, пробка, первичная кора); 2—S — луб: 2 — лубяные волокна, з — ситовидные трубки, 4 — клетки-спутники (3 и 4 вынесены на поля, где изображены более крупно), 5 — клетки лубяной паренхимы; 6 — клетки камбия, в крайних слоях растянутые, дифференцирующиеся; 7—9 клеточные элементы древесины: 7 — клетки сосудов, 8 — древесные волокна, д — клетки древесной паренхимы (7, 8 ш 9 показаны также крупно); 10 — клетки сердцевины.
99. Что общего и в чем различие анатомического строения плоского листа и хвои?
Плоский лист покрыт со всех сторон эпидермой (кожицей). В обычных, горизонтально ориентированных листьях верхний и нижний эпидермис различаются. В целом, кожица состоит из тесно сомкнутых паренхимных клеток с извилистыми очертаниями. Клетки нижнего эпидермиса обычно более мелкие и более извилистые.
В верхнем эпидермисе в типичном случае сильнее развиты кутикулярные слои, однако опушение приурочено главным образом к нижней стороне листа. Здесь же, в нижнем эпидермисе обычно располагаются и устьица.
Типичный эпидермис однослоен. Под эпидермисом залегает основная зеленая хлорофиллоносная ткань - мезофилл. У большинства растений хлоренхима дифференцирована на столбчатую (палисадную) и рыхлую (губчатую) ткани. В типичном случае к верхней кожице примыкает столбчатый мезофилл, к нижней - губчатый.
Нередко эти две ткани соединяются посредством особых собирательных (воронковидных) клеток. Характерным элементом многих листьев являются склереиды, придающие нежным листовым пластинкам дополнительную прочность.
Не менее свойственны листьям крупные клетки - идиобласты с отбросами, например, кристаллами солей (как цистолиты у фикуса). В листьях они имеют особое значение. Так, у некоторых галофитов, растений, произрастающих на засоленных субстратах, единственный способ вывести избыток солей из организма - опадание листьев.
Проводящая система в листе представлена сосудисто-волокнистыми коллатеральными закрытыми пучками. Характерно расположение проводящих тканей: ксилема располагается в верхней части пучков, флоэма - в нижней.
Именно по этому признаку можно точно определить верх и низ листовой пластинки. Все другие признаки изменчивы, так, устьица могут находиться сверху и т. д.
Хвоя - листья многих хвойных деревьев и кустарников. Обычно игольчатой или чешуевидной формы, хорошо приспособлена к неблагоприятным условиям жаркого лета, холодной зимы. Хвоя обычно многолетняя, опадает постепенно. Эпидермис хвои с толстой кутикулой, устьица погруженные. Мякоть листа состоит из паренхимных клеток (с хлорофилловыми зёрнами), стенки которых у сосны образуют внутри клетки выросты и складки, увеличивающие фотосинтезирующую поверхность хвои. В центре хвои расположены 1—2 проводящих пучка. Хвоя многих видов содержит смоляные ходы. Живая хвоя выделяет в атмосферу особые вещества — фитонциды, влияющие на состав микрофлоры воздуха. Опавшая хвоя образует медленно перегнивающую лесную подстилку.
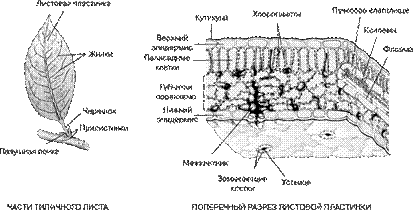
Рис. 6. Строение плоского листа
Список использованной литературы
1. Алексеев Ю.А., Вахромеева М.Г., Денисова Л.В., Никитина СВ. Лесные травянистые растения. М.: Агропромиздат, 1988.
2. Власова Н.П. Практикум по лесным травам. М.: Агропромиздат, 1986.
3. Васильев А.Е., Воронин Н.С., Еленевский А.Г. и др. Ботаника. Морфология и анатомия растений. М.:
Просвещение,1998.
4. Грин Н., Стаут У., Тейлор Д. Биология. Т. 1-3. М.: Мир, 1990.
5. Гэлстон А., Девис П., Сэттер Р. Жизнь зеленого растения. М.: Мир, 1983.
6. Жизнь растений. Т.1-6 М.: Просвещение, 1980.
7. Крамер П., Козловский Т. Физиология древесных растений. М.: Лесная промышленность, 1983.
8. Матвеева Г.В., Тарабрин А.Д. Ботаника. М.:Агропромиздат, 1989.
9. Полевой В.В. Физиология растений. М.: Высшая школа, 1989.
10. Родионова А.С, Барчукова М.В. Ботаника. М.:Агропромиздат, 1990.
11. Родионова А.С. Лесная ботаника. М.: Лесная промышленность, 1980.
12. Рейвн П., Эверт Р., Айкхорн С. Современная ботаника. Т.1-2. М.: Мир, 1990.
13. Сибирякова М.Д. Типы леса лесорастительных районов Европейской части СССР с иллюстрацией подлесной флоры. М.: Гослесбумиздат, 1962.
14. Хржановский В.Г., Пономаренко С.Ф. Ботаника. М.: Агропромиздат, 1988.