
The Blind Watchmaker
| Вид материала | Документы |
(1991)
Компьютерные программы, и эволюция самой эволюции
Компьютерная программа “Биоморф”, описанная в третьей главе сейчас доступна для “Эппл Макинтош”, “Нимбус” и IBM-совместимых компьютеров. Все три программы поддерживают девять основных "генов", необходимых для генерации биоморфов, как показанных в третьей главе, так и триллионов им подобных (или не подобных). Версия программы для компьютера “Макинтош” также поддерживает ряд дополнительных генов, производя "сегментированные" биоморфы (с "градиентной" сегментацией) и биоморфы, обладающие различными видами симметрии. Эти усовершенствования биоморфных хромосом, вместе с новой цветной версией программы, сейчас уже разработанные для “Макинтош-2”, но ещё не выпущенные, навели меня на мысли о "эволюции самой эволюции". Это новое издание “Слепого часовщика” предоставляет мне возможность поделиться некоторыми из этих мыслей.
Естественный отбор может работать лишь с теми вариациям, которые сгенерированы мутациям. Мутация определяется как "случайная", но это означает лишь то, что она не смещена систематически к усовершенствованию, и мы можем постичь лишь в высшей степени неслучайную субпопуляция всех вариаций. Мутация действует, изменяя процесс уже действующего эмбриогенеза. Вы не сможете сделать слона с помощью мутации, если действующий эмбриогенез - эмбриогенез осьминога. Это вполне очевидно. Что же было мне менее очевидно, пока я не начал играть с усовершенствованной программой Слепого часовщика, так это то, что не всякий эмбриогенез одинаково плодородно-отзывчив на попытки побудить его к дальнейшей эволюции.
Представьте себе, что внезапно открылись широкие перспективы эволюционных возможностей - скажем, целый континент опустел вследствие природной катастрофы. Какие виды животных заполнят эволюционный вакуум? Конечно, это должны быть потомки индивидуумов, успешных в деле выживания в пост-катастрофических условиях. Но ещё интереснее, что некоторые виды эмбриогенеза могут быть особенно удачны не только для выживания, но и для эволюции. Возможно, что причина, по которой млекопитающие так распространились после вымирания динозавров, состояла не только в том, что млекопитающие были успешны в индивидуальном выживании в пост-динозавровом мире. Возможно, что способ развития тел млекопитающих оказался также "хорош" и для генерации большого разнообразия типов – плотоядные, травоядные, муравьеды, лазающие, роющие, плавающие, и так далее, в силу чего про млекопитающих можно сказать, что они хороши и в эволюции.
Какое отношение это имеет к компьютерным биоморфам? Вскоре после разработки программы Слепого часовщика, я экспериментировал с другими компьютерными программами, которые делали то же самое, однако использовали другой эмбриологический базис – другие принципы рисования тел, на которые могли воздействовать мутация и отбор. Эти другие программы, будучи поверхностно подобными "Слепому часовщику", оказались уныло скудными по предлагаемой широте эволюционных возможностей. Эволюция постоянно застревала в бесплодных тупиках. Вырождение, казалось, было самым типичным результатом даже тщательнейше ведомой эволюции. Напротив, эмбриология ветвящихся деревьев, лежащая в основе программы Слепого часовщика, выглядела “постоянно беременной” возобновляемыми эволюционными ресурсами; не наблюдалось никакой тенденции к автоматическому вырождению в ходе эволюции – богатство, многогранность, и даже красота, казалось, бесконечно возрождались по мере вырисовывания поколений.
Однако, как бы плодовитой и разнообразной ни была биоморфная фауна, порождаемая оригинальной программой "Слепого часовщика", я постоянно наталкивался на явные барьеры на пути дальнейшей эволюции. Но раз эмбриология “Слепого часовщика” эволюционно столь превосходит альтернативные программы, то нельзя ли предложить какую-нибудь её модификацию, расширяющую эмбриологические правила вычерчивания, которая могла бы сделать “Слепого часовщика” ещё более обильным в смысле эволюционного разнообразия? Или, другими словами, нельзя ли основную хромосому из девяти генов расширить в направлении большей плодотворности?
Разрабатывая оригинальную программу "Слепого часовщика", я преднамеренно старался не использовать мои биологические знания. Моя цель состояла в демонстрации мощи неслучайного отбора случайной вариации. Я хотел увидеть биологию, совершенство и красоту как результат отбора. Я не хотел давать самому себе поводов для обвинений в том, что я сразу встроил эти качества в программу. Эмбриология ветвящихся деревьев “Слепого часовщика” была самой первой испробованной мною эмбриологией. И как оказалось, я угадал, что стало ясно по результатам моего последующего разочаровывающего опыта с альтернативными эмбриогенезами. В своих размышлениях о способах усовершенствования основной "хромосомы", я позволил себе роскошь использовать кое-что из моих биологических знаний и интуиции. Большинство эволюционно успешных групп животных обладают сегментированным строением тела. Также в числе фундаментальных особенностей строения тела животных – виды их симметрии. Стало быть, новые гены, которые я добавил к хромосоме биоморфа, управляли сегментацией и симметрией. Мы с вами, и все позвоночные, сегментированы. Это видно по нашим рёбрам, и нашему позвоночнику, повторяющийся характер которых виден не только на самих костях, но и в связанных с ними мышцах, нервах и кровеносных сосудах. Даже наши головы в сущности сегментированы, но сегментная структура головы у взрослого незаметна никому, кроме специалистов, изучающих эмбриональную анатомию. Рыбы сегментированы более явно, чем мы (вспомните о батарее мышц, проходящих вдоль позвоночника копчёной сельди). У ракообразных, насекомых, и многоножек сегментация хорошо видна даже извне. Различие в этом отношении между многоножкой и омаром состоит в степени однородности. Многоножка подобна длинному товарному поезду с почти идентичными вагонами. Омар подобен поезду с пёстрым разнообразием товарных и пассажирских вагонов, в основе одинаковых и одинаково соединённых между собой, с придатками, исходящими из каждого. Но кое-где вагоны сварены вместе в группы, а придатки стали большими ногами или клешнями. В области хвоста вагончики и меньше, и однообразнее, а их когтистые боковые отростки стали маленьким, перообразными "лопастями".
Чтобы сделать биоморфы сегментированными, я сделал очевидную вещь - ввёл один новый ген, управляющий "числом сегментов", а также другой новый ген, управляющий "расстоянием между сегментами". Один полный биоморф старого стиля стал отдельным сегментом биоморфа нового стиля.

Выше показаны семь биоморф, отличающиеся только геном "числа сегментов”, или геном "расстояния между сегментами". Левый биоморф – наше старое знакомое ветвящееся дерево, а другие – лишь последовательно повторяющиеся вагончики из этого же дерева. Простое дерево, как все биоморфы оригинальной версии программы “Слепой часовщик, являются частным случаем "односегментного животного".
Пока что я говорил только об однородной сегментации, как у многоножек. Сегменты у омара отличаются друг от друга сложным образом. Простейший стиль изменения сегментов – по "градиентам". У мокрицы сегменты больше похожи друг на друга, чем у омара, но всё же они не так однородны, как у типичной многоножки (фактически некоторые с виду "мокрицы" являются многоножками). Мокрицы уже спереди и сзади, и шире в середине. При отображении её по мере движения от головы к хвосту, градиент размера сегментов будет максимальным в середине. Другие сегментированные животные, например вымершие трилобиты, имеют наибольшую ширину спереди, и заострение сзади. У них более простой градиент размера, достигающий максимума с одного конца. Именно этому более простому виду градиента я стремился подражать в своих сегментированных биоморфах. Я реализовывал это, прибавляя константу (возможно - отрицательную) к значению конкретного гена, идя от головы к хвосту. Из следующих трёх биоморфов, у левого нет никаких градиентов, у среднего есть градиент гена 1, а у правого – гена 4.

Нарастив основную хромосому биоморфа этими двумя генами, и ассоциированными генами градиента, я был готов дать волю новому стилю биоморфной эмбриологии в компьютере, и смотреть, что он может делать в ходе эволюции. Сравните следующую картину с рисунком 5 третьей главы, где у всех биоморф отсутствует сегментация.
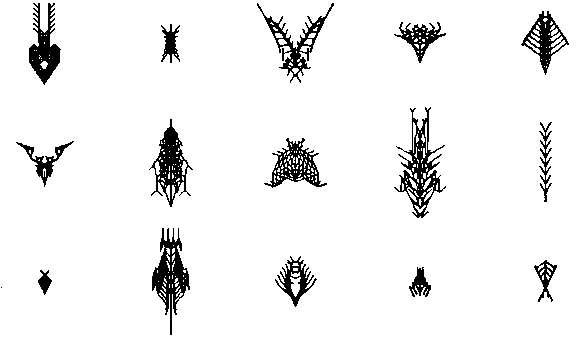
Думаю, вы согласитесь, что стал доступен более "биологически интересный" диапазон эволюционного многообразия. "Изобретение" сегментации, как нового крупного достижения эмбриологии, выпустило на волю новый эволюционный потенциал в стране компьютерных биоморфов. Я догадываюсь, что нечто подобное случилось в эволюции позвоночных, первых сегментированных предков насекомых, омаров и многоножек. Изобретение сегментации было поворотным событием эволюции.
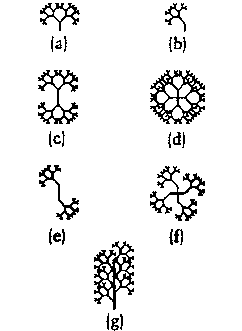
Симметрия была другим очевидным новшеством. Оригинальные биоморфы “Слепого часовщика”, были ограничены вертикально-осевой симметрией. Я добавил новый ген, сделавший симметрию не обязательной. Этот новый ген определял, будет ли биоморф с его оригинальным набором девяти генов напоминать базовое дерево (a) или (b). Другие гены определяли, будет ли он симметрично отражён сверху вниз, (c), или обладать полной четырёхсторонней симметрией, (d). Эти новые гены могли изменяться в любой комбинации, как в (e) и (f). Я добавил подсказанное растениями ограничение: когда сегментированные животные были асимметричны относительно горизонтальной оси, то противостоящие сегменты должны быть асимметричны в противостоящих направлениях, как в (g).
Вооруженный этими новыми генами, я снова приступил к энергичной программе разведения животных, чтобы посмотреть, может ли эта ещё более новая эмбриология порождать ещё обильную эволюцию. Вот набор сегментированных биоморф с асимметрией по средней линии:

А здесь приведено несколько радиально-симметричных биоморф, сегментация которых, если она вообще есть, может быть столь же скрытной, как и у головы взрослого человека:

Ген полной радиальной симметрии более соблазняет селекционера на разведение приятных абстрактных проектов, нежели биологически реалистичных, которые я искал изначально. Это даже более актуально для цветной версии программы, которую я сейчас разрабатываю.
Одна группа животных - иглокожие (включающая морскую звезду, морского ёжа, офиура и морскую лилию), очень нетипичны своей пятисторонней симметрией. Я не сомневаюсь, что как бы упорно я, или кто-нибудь другой ни пытались сделать это, но мы никогда не получили бы пятилучевую симметрию в результате случайной мутации существующего эмбриогенеза. Это бы потребовало новой "переломной" новации в эмбриологии биоморф, так что я и не пытался. Но в природе иногда бывают уродливые морские звёзда и морские ёжи с четырьмя или шестью конечностями вместо обычных пяти. Исследуя Страну биоморф, я натолкнулся на поверхностно схожие с морской звездой или ежом формами, которые подтолкнули меня на селекцию в сторону увеличения сходства. Вот коллекция биоморф, сходных с иглокожими, хотя ни один из них не обладает необходимым для полного подобия пятью конечностями:

В качестве финального испытания многогранности моей новой эмбриологии биоморф, я поставил себе задачу размножения биоморфного алфавита, достаточно полного, чтобы написать своё имя. Каждый раз, когда я наталкивался на биоморф, который был похож, пусть и слегка, на букву алфавита, я селектировал его, дабы увеличить сходство. Приговор этой честолюбивой попытке неоднозначен, мягко говоря. "T" и "N" почти точны. "A" и "H" респектабельны, разве что чуть нескладны. "D" плоха, а вот вывести должную "K", я подозреваю, просто невозможно - я был вынужден слукавить, заимствуя вертикальный штрих у "W". Подозреваю, что нужно было бы добавить ещё один ген, прежде чем правдоподобная "K" могла бы быть развита.
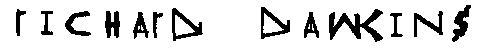
После этой несколько неграмотной попытки подписать своё имя, я имел больше везения с эволюцией имени того вдохновляющего артефакта, с помощью которого вся эта работа и была проделана
На меня произвело сильное впечатление, надеюсь, поддержанное этими иллюстрациями, то, как введение нескольких радикальных перемен в фундаментальной эмбриологии биоморф открыло новые перспективы эволюционных возможностей, которые были просто недоступны первоначальной программе, описанной в третьей главе. И как я сказал выше, я полагаю, что нечто подобное происходило при различных обстоятельствах в эволюции некоторых известных групп животных и растений. Изобретение сегментации нашими предками, и независимо – предками членистоногих, вероятно является только одним примером нескольких переломных событий нашей эволюционной истории. Эти переломные события, по крайней мере - если на них смотреть с мудростью ретроспективы, отличаются от обычных эволюционных изменений. Наши первые сегментированные предки, и первые сегментированные предки дождевых червей и насекомых, возможно и не были особенно хороши в выживании как индивидуумы, хотя очевидно они всё же выживали; в противном случае нас, их потомком, здесь бы не было. Моя позиция сейчас такова, что изобретение сегментации этими прародителям было значимее, чем одна лишь новая техника выживания, такая как более острые зубы или более острое зрение. Когда сегментация добавилась к эмбриогенетическим процедурам наших прародителей, то безотносительно к тому, помогло ли это им в выживании, или не очень, но линии, которым они принадлежали, внезапно стали лучше эволюционировать.
Современные животные – мы, позвоночные и все наши попутчики на этой планете, наследуют гены непрерывной линии прародителей, которые были удачливы в индивидуальном выживании. Это я старался максимально ясно дать понять в “Слепом Часовщике”. Но мы также наследуем те эмбриогенетические процедуры прародительских линий, которые были хороши для эволюции. Существовал своего рода отбор более высокого уровня среди линий, не в пользу их способности выживать, а в пользу их способности долго эволюционировать. Мы носим накопленные усовершенствования множества переломных событий, и изобретение сегментации - только одно из них. В направлении улучшений развивались не только тела и поведение. Мы можем даже говорить, что сама эволюция эволюционировала. Имела место прогрессивная эволюция способности к эволюции.
Версия программы Слепой часовщика для Макинтоша имеет меню опций, позволяющее включить или выключить главные категории мутаций. Выключив все новые типы мутаций, мы возвращаемся к более ранней версии программы (или существующей IBM версии). Проведите селекцию какое-то время при этих условиях, и вы получите некоторое впечатление не только огромности диапазона фаун, разрешённых в ранней программе, но и её ограничений. И если вы затем включите, скажем, мутации сегментации, или мутации симметрии (или если вы переключаете с IBM на Макинтош), вы можете испытать ликующее чувство, в чём-то подобное чувству освобождения, которое, возможно, сопровождало великие переломные события эволюции.