
|
< Предыдущая |
Оглавление |
Следующая > |
|---|
2.2. Механизмы образования ассоциативных связей
Способность к формированию ассоциативных условно-рефлекторных реакций выявляется у живых организмов на уровне механизмов функционирования участков нейронных сетей. Процесс образования ассоциативных связей наглядно продемонстрирован в опытах на беспозвоночных животных, в частности на морском брюхоногом моллюске аплизии (Aplysia punctata). Большие по размеру нейроны беспозвоночных (порядка 100 мкм) позволяют проводить эксперименты по их индивидуальному раздражению под визуальным контролем.
Схема ассоциативного обучения в этих экспериментах имела следующий вид. В ходе своего нормального поведения аплизия отвечает слабым втягиванием жабры в ответ на слабое прикосновение к сифону. (Такое воздействие может рассматриваться как аналог условного сигнала, если исходить из предположения, что околопороговая сила раздражения связана с практическим отсутствием ответа.) С другой стороны, в ответ на сильный удар током по
"голове" аплизия реагирует сильным втягиванием жабры (что можно рассматривать как аналог безусловного рефлекса). После ряда совместных воздействий животное реагирует на слабое прикосновение ксифону таким же активным втягиванием жабры, как и в ответ на удар током по "голове".
Упрошенная схема участка нейронной сети, реализующей такое поведение, приведена на рис. 4.4. На рис. 4.4, А представлен вариант, при котором ассоциативное обучение реализуется на уровне контактов нейронов трех типов: нейрона - датчика касаний сифона (/), нейрона - датчика болевого воздействия (3) и нейрона, запускающего двигательный акт сжатия мышцы мантии (2). На рис. 4.4, Б рассмотрен более сложный вариант, при котором в схему включен дополнительный элемент - интернейрон (4), играющий роль прообраза участка нейронной сети, обладающего неко-
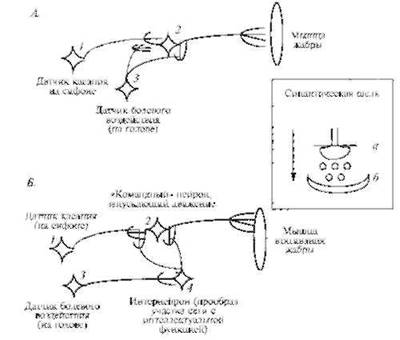
Р и с. 4.4. Схема двух вариантов образования условного рефлекса (ассоциативной связи) в участке нейронной сети морской улитки аплизии (Aplisia punctata). В обоих вариантах цепь безусловного рефлекса:
раздражение датчика боли (нейрон 3) - сжатие мышцы (нейрон 2). А - упрошенный вариант: 6 - вариант с использованием вставочных нейронов (интернейронов). На вставке показана схема синаптического соединения, где (а) прс-.а (б) постеинаптическис окончания нейронов, кружки - молекулы нейромедиатора. стрелка указывает направление прохождения сигнала
торой интеллектуальной функцией. Группы интернейронов или вставочных нейронов, то есть нейронов, вставленных между сенсорными и двигательными нейронами, действительно становятся все более и более сильно развитыми по мере продвижения животных по филогенетической лестнице.
Схема первого упрошенного варианта формирования цепи условного рефлекса такова. Вначале касание сифона не ведет к реакции втягивания жабры, но после нескольких десятков сочетаний касания сифона и последующего удара током по голове появляется условный рефлекс. Механизм рефлекса заключается в том, что активация нейрона 3 облегчает проведение сигнала через синапсы между нейронами / и 2.
Во втором варианте механизм рефлекса заключается в активации интернейрона 4, окончания которого увеличивают локальный электрический потенциал в определенном районе "командного" нейрона 2. В итоге этого достаточно сложного процесса происходит изменение геометрии в области синаптических соединений отростков интернейрона 4, командного нейрона 2 и нейрона-датчика касаний /. ^го изменение связано с увеличением размеров так называемых "шипиков", что и представляет собой материальную реализацию процесса обучения.
Места соединения нейронов - синапсы, или синаптические щели, - играют важную роль в механизмах обучения. Структуру этих щелей образуют пре- и постсинаптические нейронные мембраны, разделенные межнейронным пространством. Нервный импульс (или, как его часто называют, потенциал действия) передается от пресинаптической мембраны одного нейрона к постсинаптической мембране другого.
При этом сам процесс передачи представляет собой сложное, многоэтапное событие, реализуемое посредством работы электрических и молекулярных механизмов. В результате прохождения нервных импульсов по "входному", пресинаптическому нейрону в районе синаптической щели из специализированных структур пресинаптических окончаний выделяются молекулы нейромедиатора, которые пересекают синаптическую щель, активируют рецепторы постсинаптической мембраны и вызывают появление по-стсинаптических потенциалов, рост "шипиков", изменение геометрии участков сети и в конечном счете запоминание и ассоциативное обучение.
На основании экспериментальных данных последних десятилетий становится ясно, что материальным субстратом закрепления ассоциативных связей, в частности и запоминания вообще.
являются именно синаптические структуры. В результате образования ассоциации, запоминания или фиксации любой информации происходит формирование новой конфигурации нейронных связей участков нейронных сетей. Сетевые структуры формируют сязасчеттого, что отела каждого нейрона (нервной клетки)отходит множество веточек дендритов, получающих входные сигналы от других клеток, и один тонкий ветвящийся аксон, по которому сигнал после обработки передается к другим нервным клеткам. При таком строении за счет своего ветвящегося дендритного дерева каждый нейрон может получать сигналы от множества других нервных клеток. Обработанный сигнал также может быть передан многим нейронам за счет ветвящихся концевых окончаний аксона.
Интересно отметить, что структура нейронных сетей впервые стала доступна для наблюдения только в конце XIX века, когда анатом из Милана Камилло Гольджи почти что неожиданно для себя открыл метод серебрения. Метод позволял окрашивать небольшие участки нервной ткани, причем отдельные элементы выделялись настолько четко, что были видны мелкие детали клеточного тела, дендритов и аксонных окончаний. Не менее интересно, что сам Гольджи, будучи уже нобелевским лауреатом, не верил в существование отдельных нейронов и представлял нервную ткань как непрерывную сеть. Взгляд на нейронную сеть как на структуру, состоящую из отдельных нервных клеток и разделяющих их синаптических образований, по-видимому, впервые высказал современник Гольджи, тоже нобелевский лауреат нейроанатом Рамон-и-Кахал. Как пишут, Гольджи не верил до юлам Кахала и отказывался даже разговариватьсним (50). Воистину, смотрят все, но видят немногие, хотя, с другой стороны, немногим же дано воспользоваться случайностью и использовать предоставленную им возможность.
Структурные изменения при формировании нейронных сетей, реализующих ассоциативные связи, происходят на уровне мембран ветвящихся пресинаптических окончаний (аксонов) одного нейрона (выходных элементов нервной клетки) и ветвящихся посте и найти чески X окончаний (дендритов) другого нейрона (входных элементов нервной клетки). В итоге этого процесса происходит изменение геометрии в областях дендритных и аксонных окончаний. Эти изменения связаны, в частности, с увеличением количества особых выростов дендритов, так называемых "шипиков", а также с изменением размеров этих "шипиков". В свою очередь, изменения геометрии ведут к изменениям электрических свойств межнейронных контактов, то есть к изменению проводимости и возбудимости определенных нервных связей. По-видимому, именно таким путем реализуется механизм "проторения" нового пути на участке нейронной сети.
В настоящее время накоплено определенное количество данных о молекулярных механизмах этих процессов. Начальные этапы этих процессов связаны с тем, что высвобождающиеся из пресинаптического окончания молекулы ней роме диатора воздействуют на рецептор постсинаптической мембраны. В результате происходит целый каскад процессов, связанных с открытием или закрытием ионных каналов, что приводит к кратковременным эффектам изменения электрической активности и, следовательно, являет собой молекулярную основу кратковременной памяти (2: III, 333).
С другой стороны, получен ряд данных, позволяющих рассматривать гипотезы о молекулярных основах долговременной памяти, обеспечивающей изменение геометрии синаптических структур и проторение новых путей в нейронных сетях. Полагают, что нейромедиатор в процессе взаимодействия с рецепторами постсинаптической мембраны запускает особый цикл молекулярных процессов, что приводит в конечном счете к экспрессии различных групп генов, расположенных в ядрах активированных в данном процессе нервных клеток. Некоторые из этих генов известны и характерны тем, что становятся активными в клетках, проводящих синтез белковых молекул, которые транспортируются к соответствующим местам мембраны, включаются в нее и изменяют в конечном счете ее форму и размеры (2: III, 333-334; 49:245-352).
Роль структурных генов в механизмах ассоциативного запоминания активно изучается на классических объектах генетики - плодовых мушках Drosophila. Например, дрозофил можно научить избегать любой запах, если этот запах будет неоднократно сопровождаться ударом тока. При этом можно выделить тех "бестолковых" мушек, которые либо неспособны к образованию такой ассоциации, либо очень быстро забывают ее. Оказывается, что такие мушки являются мутантами, у которых повреждены или отсутствуют разные группы генов.
|
< Предыдущая |
Оглавление |
Следующая > |
|---|