
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
ЭКОЛОГО-БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПРОСА (Panicum miliaceum L.) И ТРАДИЦИИ ЕГО ВОЗДЕЛЫВАНИЯ В ЦЕНТРАЛЬНОЙ ТЫВЕ
Автореферат кандидатской диссертации
На правах рукописи
МОНГУШ Лаида Кара-ооловна
ЭКОЛОГО-БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ПРОСА (Panicummiliaceum L.) И ТРАДИЦИИ
ЕГО ВОЗДЕЛЫВАНИЯ В ЦЕНТРАЛЬНОЙ ТЫВЕ
03.02.01 - ботаника (биологические науки)
АВТОРЕФЕРАТ
диссертации на соискание учёной степени
кандидата биологических наук
а а Улан-Удэ - 2012
Работа выполнена на кафедре общей биологии ФГБУО ВПО
Тывинский государственный университет (г. Кызыл)
Научный руководитель: доктор биологических наук, профессор
а Намзалов Бимба Батомункуевич
Официальные оппоненты: доктор биологических наук, профессор
ФГБОУ ВПО Алтайский
государственный университет
Терехина Татьяна Александровна
а кандидат биологических наук,
ст. научный сотрудник ФГБНУ
Институт общей и экспериментальной
биологии
а Бадмаева Наталья Карловна
а
Ведущая организация: Федеральное государственное
бюджетное научное учреждение
Тувинский институт
комплексного освоения
природных ресурсов СО РАН
Защита состоится л21 мая 2012 г. в 16.00 часов на заседании диссертационного совета Д 212.022.03 при Федеральном государственном бюджетном образовательном учреждении высшего профессионального образования Бурятский государственный ауниверситет по адресу: 670000,
г. Улан-Удэ, ул. Смолина, 24а, конференц-зал.
Факс: (3012) 210588, e-mail: ;
аС диссертацией можно ознакомиться в библиотеке ФГБОУ
Бурятский государственный университет (670000, г. Улан-Удэ,
ул. Смолина, 24а)
Автореферат разослан л 20 апреля 2012 г.
Учёный секретарь
диссертационного совета,
канд. биол. наук ааН.А. Шорноева
а
ВВЕДЕНИЕ
Актуальность темы. Злаковые растения, несомненно, являются первостепенными по значимости в жизнеобеспечении человека. Просо, также как и ячмень, относится к древнейшим пищевым растениям, введенным в культуру (Жуковский, 1964).
В Туве в условиях экстраконтинентального климата и исключительного разнообразия экотопов в контрастных ландшафтах межгорных котловин многообразие дикорастущих и культивируемых форм проса наиболее велико. Следует отметить, что тувинский этнос сохранил сорта местной селекции проса, которые представляют непреходящую ценность в современном пищевом рационе. Как известно, Центральная Азия относится к одному из центров происхождения культуры проса (Вавилов, 1929). Однако в последние 2-3 десятилетия просо в Тыве практически не высевается, если не считать частные семейно-родовые хозяйства, которые еще продолжают традиции его возделывания.
аВ связи с этим становятся наиболее актуальными исследования эколого-биологических особенностей тувинской популяции проса, изучение опыта традиционного его возделывания, пищевых свойств этой культуры. Результаты исследования позволят существенно облегчить работу по сохранению их генофонда. Решение этих вопросов касается реализации ключевой проблемы современности - сохранения биоразнообразия естественной и культурной флоры планеты.
Цель исследования. Цель исследования - выявить эколого-биологичес-кие особенности проса (Panicummiliaceum L.) в Центральной Туве, а также дать анализ его традиционного возделывания в регионе.
Задачи исследования:
- Дать характеристику природно-климатических условий района исследования.
- Выявить ценофлору сорняков в посевах проса и провести анализ ее структуры (таксономической, эколого-географической и биоэкологической).
- Установить разнообразие просовых сообществ, а также выявить особенности их эколого-ценотической структуры.
- Изучить эколого-биологические особенности проса, а также дать морфобиологические характеристики его экотипов.
- Рассмотреть особенности традиционного возделывания проса в Тыве, определить перспективные экотипы для селекции.
Защищаемые положения.
1. Разнообразие сорняков в посевах проса в Центральной Тыве насчитывает 41 вид сосудистых растений (31 род и 19 семейств), где преобладают виды семейств Asteracea, RosaceaиPoaceae. Это травянистые моно- и поликарпики с широкими голарктическим и евразийским ареалами, мезоксерофитной экологии, что в целом характерно для флоры сорно-полевой растительности.
2. Эколого-биологические исследования чаа-хольской популяции проса выявили в ее составе формирование двух экотипов со специфическими морфологическими признаками вегетативных и генеративных органов, анатомического строения листьев, а также особенностями ритмики развития и продуктивности, которые будут полезны в селекционной работе с местными популяциями проса.
3. Анализ этнических традиций культуры проса в Тыве выявил разнообразие форм в местных популяциях вида, нам удалось раскрыть особенности традиционных форм возделывания проса, уборки и хранения урожая, способов приготовления из него пищи. Это является ценным источником новых знаний об одном из древнейших пищевых растений.
Научная новизна и практическая значимость работы.
Впервые дана обстоятельная флористическая и эколого-биологическая, фитоценотическая характеристика агроценозов проса в Центральной Тыве. Ценофлору посевов проса слагает 41 вид из 31 рода и 19 семейств.
Проведён анализ структуры флоры в систематическом, эколого-фитоценотическом и географическом аспектах. Систематический спектр флоры показал значительное участие видов семейств Rosaceae, Asteraceae и Poaceae. Остальные семейства представлены преимущественно монотипными родами. В эколого-географическом отношении преобладают мезоксерофиты широкого евразийского распространения.
Анализ разнообразия просовых сообществ региона с использованием эколого-фитоценотического подхода позволил осуществить их классификацию, в результате выделено 3 ассоциации. На примере полынно-сурепково-просового агроценоза изучены важнейшие эколого-ценотические особенности сообщества.
Отмеченные в чаа-хольской популяции проса два экотипа имеют морфологические отличия по признакам вегетативных и генеративных органов, анатомического строения листьев, а также по феноритмике развития и показателям всхожести семян. Выявленные признаки представляют интерес в селекционной работе проса в Тыве.
Впервые получены ценные этноботанические сведения по традициям возделывания проса в Тыве. Описаны особенности технологии выращивания этой культуры, ухода, хранения и сбора урожая, а также изучены национальные традиции приготовления пищи из проса. Выявленные экотипы проса составляют не только ценный материал в практике его возделывания в регионе, они важны для сохранения генофонда культурной флоры.
Результаты исследований могут быть использованы при подготовке спецкурсов по экологии и систематике культурных растений и ботанической географии в вузах.
Материалы и методы.
В работе обобщен материал исследований в период с 2009 по 2011 г. В ходе полевых работ изучено тринадцать ценопопуляций в Чаа-Хольском и Сут-Хольском кожуунах Республики Тыва с использованием маршрутных и полустационарных методов, собрано около 250 листов гербария, выполнено 13 геоботанических описаний просовых агроценозов.
Маршрутные и полустационарные исследования проводились посредством общепринятых флористических и геоботанических методов. При полустационарных исследованиях использованы методики: фенологические наблюдения по И.Н. Бейдеман (1960), И.В. Борисовой (1961), при определении фитомассы - метод укосных квадратов, с площадок 1х1 м, в 3-5-кратной повторности.
Для выявления эколого-ценотических особенностей эталонного полынно-сурепково-просового агроценоза были заложены серии 0,5 м трансектов с регистрацией всех видов, всего было отмечено 700 случаев пересечения особей видов растений. Для выявления сопряженных связей видов использовался метод корреляционных плеяд П.В. Терентьева (1960).
Анатомическую структуру листа изучали общепринятым методом (Дженсен, 1965; Пронзина, 1960), на фиксированных образцах, собранных в период бутонизации. Поперечные срезы делали вручную с помощью бритвы - временные препараты. Просмотр срезов проводили методом световой микроскопии (лAmpleval Germany, DDR). Размеры и число устьиц определяли по методу Т.Н. Годнева и Г.А. Липской (1965) и выражали в мкм. Рисунки сделаны на микроскопе TM-1000 Tabletop Microscope (HITACHI).
Проращивание семян проса для определения энергии прорастания и всхожести проводилось по ГОСТу 13056.6-97 с соблюдением всех требований. Статистическая обработка полученных данных проводилась с использованием пакета программ Excel (Statistica 7.0).
Для изучения видового состава растений использовали: Определитель растений Тувинской АССР (2007) и Флора Сибири (1987-1997).
Апробация работы. Основные результаты диссертационной работы доложены на VIII Междунар. науч.-практ. конф. Проблемы ботаники Южной Сибири и Монголии (Барнаул, 2009); I Межрегион. науч.-практ. конф. для молодых ученых, аспирантов и студентов Актуальные проблемы исследования этноэкологических и этнокультурных традиций народов Саяно-Алтая (Кызыл, 2009); междунар. науч. конф: чтения памяти проф. М.М. Кожова Проблемы экологии (Иркутск, 2010); IX Санкт-Петербург. этнограф. чтения Традиционное хозяйство в системе этноса (Санкт-Петербург, 2010); всероссийской науч.-практ. конф. Современные проблемы этноэкологии и традиционного природопользования (Улан-Удэ, 2010) и Первой междунар. науч. конф. Сорные растения в изменяющемся мире: актуальные вопросы изучения разнообразия, происхождения, эволюции (Санкт-Петербург, 2011).
Кроме этого, материалы диссертации докладывались на заседаниях кафедры экологии и общей биологии ТывГУ (2009, 2010, 2011).
аПубликации. По материалам диссертации опубликовано 9 работ, из них две статьи в рецензируемых изданиях.
Структура и объём работы. Диссертация состоит из введения, 5 глав, выводов, литературы, включающей 132 наименования и приложения. Работа изложена на 137 с. машинописного текста, включает 16 таблиц и 21 рисунок.
ГЛАВА 1. Природные условия Центральной Тувы. Рельеф межгорных котловин Тувы в разные годы изучали С.В. Обручев (1942), С.Л. Кушев (1957), Б.Н. Лиханов (1956), Г.А. Чернов (1988) и др. В центральной части Тувы располагаются обширная Центрально-Тувинская котловина (рис. 1), Хемчикская, Улуг-Хемская и Турано-Уюкская межгорные впадины (Кушев, 1957). В Хемчикской котловине, в приустьевой части р. Хемчик, обособляется геоморфологически четко выраженная депрессия - Чаа-Хольская.
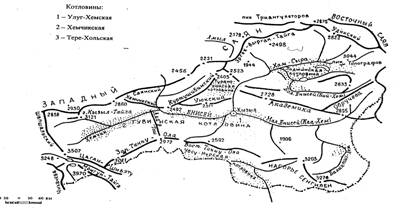
Рис. 1. Орографическая схема Центрально-Тувинской котловины
аКлимат района исследования - резко континентальный с холодной и малоснежной зимой, жарким летом с большой амплитудой абсолютных и суточных температур (Ефимцев, 1957). Среднегодовая температура воздуха - -5-7С, абсолютный максимум приходится на июль +36-39С. Среднегодовое количество осадков 200-230 мм.
В разнообразии почвенного покрова основной фон составляют почвы каштанового ряда в центральных частях котловины (абсолютные высотные отметки - 700-900 м.). Это каштановые почвы в сочетании со светло-каштановыми супесчаными на подгорных шлейфах (Носин, 1963). Фрагментарно распространены черноземы в теплых и более увлажненных распадках предгорий (900-1200 м.). По долинам рек на террасах преобладают лугово-дерновые суглинистые и в прирусловых частях - пойменные аллювиальные почвы.
ГЛАВА 2. Ареал, центр происхождения и особенности биологии проса (Panicum miliaceum L.). Во флоре Сибири род Panicum L. представлен одним культурным видом Просо посевное или обыкновенное (P. miliaceum L.), а также сорно-полевой его разновидностью - P. miliaceumsubsp. ruderale (Kitag.) Tzvelev. Дикая предковая форма вида не известна, это характерно для тех растений, которые давно, на заре человечества, введены в культуру. Н.И. Вавилов (1929) центром происхождения проса считает районы Восточной и Центральной Азии, где заключен полностью весь расовый и разновидностный состав линнеевского вида Panicummiliaceum (с. 51). Несколько шире рассматривают предковый ареал вида Р.В. Камелин (2005) и И.Г. Чухина (2007), которые включают, кроме того, территории Северного Китая и Средней Азии на основе индийского P. mileare Lam.
Просо обыкновенное - морфологически весьма вариабельное однолетнее растение с метельчатым и более компактным кистевидным соцветием. Колоски двуцветковые, один из которых тычиночный или недоразвитый, представлен нижней колосковой чешуей. Колосковые чешуи обоеполого цветка одинаковые, с вытянутой верхушкой. Созревшие колоски не отваливаются; зерновки пленчатые, голые, блестящие и разнообразные, по цветовым оттенкам от желтого до темно-бурого и фиолетового (Талиев,1931). К числу важнейших систематических признаков относятся: форма метелки, плотность ее, наличие бугорков у основания веточек, окраска созревших колосковых чешуй, высота растения, а также опушенность листьев, размеры семян и продолжительность вегетационного периода. Очень интересны наблюдения Н.И. Вавилова (1929), касающиеся проблемы географического центра проса. Так, им отмечается исключительное внутрипопуляционное разнообразие проса в Монголии, особенно оно проявляется в цвете семян, форме метелок и вегетационному периоде. Вероятно, это вполне характерно и для тувинских образцов проса.
Экологически просо хорошо выносит засуху - почвенную и воздушную, относится к ксерофитам. Однако на увеличение осадков, а также на орошение просо реагирует положительно, т.е. ее можно отнести к виду со скрыто патиентной стратегией (Миркин и др., 2000). Вид сильно дифференцирован на множество разновидностей (ряды региональных экотипов), в частности нами в Центральной Тыве среди чаа-хольской популяции проса выявлено два экотипа, характеристика которых дана в главе 3.
аГЛАВА 3. Сорняки просовых полей, анализ их разнообразия. Зачатки земледелия, первые опыты человечества по культивированию полезных растений, в частности злаковых культур (ячмень, проса и др.), в странах Юго-Западной Азии, Северного Китая наблюдаются не ранее 10-12,5 тыс. лет (Камелин, 2005). Вероятно, кочевые этносы Внутренней Азии, к их числу относятся верхнеенисейские урянхайцы, с их признаками очагового земледелия в дальнейшем обогатили человечество разнообразием не только пород одомашненных животных, но и богатством культурных растений. По мнению выдающегося энциклопедиста А. Гумбольдта (1936), некоторые растения сопровождали кочующего человека с одной территории на другую с отдаленнейших времен. Так, в Европе пшеница следовала за римлянами, хлопчатник - за арабами, Е (с. 63). И вероятно, эту мысль можно продолжить: просо следовало за древними тюркутами Алтая и Саян, прародителями современных тувинцев, алтайцев и западных монголов - ойратов. Это становится очевидным, если опереться на гипотезу Н.И. Вавилова (1926) о центральноазиатском происхождении культуры проса.
В связи с этим особенно важным считала Е.Н. Синская (1969) анализ сорной растительности, поскольку возможен процесс формирования культурных растений из сорняков.
а3.1. Анализ систематического, биоморфологического и экологического разнообразия сорняков. Анализ разнообразия сорняков 13 пробных площадок с посевами проса выявил 41 вид высших сосудистых растений (табл. 1). При этом большинство (10 видов из 41) относится к семейству Астровые, где преобладают полыни. Из последних к малолетним относятся Artemisiaannua, A. scopariaиA. vulgaris. Второе место занимает семейство Розоцветные и представлено только лапчатками, выделяется обилием Potentillabifurca.
Таблица 1
Семейственно-родовой спектр флоры сорняков просовых полей
№ |
Семейство Род Виды (характерные) |
Число видов |
Процент от общего числа |
1 |
Asteraceae Bercht et J. Presl. Artemisia annua L A. commutata Bess. А. glauca Pall.ex. Willd. A. scoparia Walds. S Kit. Crepis tectorum L. Taraxacum officinalis L. |
10 |
24,4 |
2 |
Rosaceae Juss. Potentilla bifurca L. P. multifida L. P. tanacetifolia Willd. еx. Schlecht |
5 |
12,2 |
3 |
Poaceae Barnhart Elytrigia repens (L.) Nevski Stipa sibirica (L.) Lam. |
4 |
9,7 |
4 |
Fabaceae Lindl. Medicago falcatа L. Melilotus albus Medicus |
4 |
9,7 |
5 |
Boraginaceae Juss. Lappula marginata (Bieb) Guerke Nonnea rossica Steven |
3 |
7.3 |
6 |
Chenopodiaceae Vent Atriplex fera (L.) Bunge Chenopodium urbicum L. |
2 |
4,8 |
4 |
Cannabaceae Martinov Cannabis sativa L. |
1 |
2,4 |
6 |
Caryophyllaceae Juss. Silene repens L. |
1 |
2,4 |
7 |
Brassicaceae Burnett Barbarea stricta Andrz. |
1 |
2,4 |
9 |
Geraniaceae Juss. Erodium stephanianum Willd. |
1 |
2,4 |
10 |
Convolvulaceae Juss. Сonvolvulus arvensis L. |
1 |
2.4 |
К группе ведущих семейств относятся Мятликовые, при этом роль Elytrigiarepens наиболее ощутима. Бобовые представлены лишь тремя видами, особенно заметно участие донника белого. Бурачниковые и маревые менее разнообразны, из этих семейств чаще отмечаются липучка и марь. Остальные семейства представлены одним видом (табл. 1). При этом необходимо отметить высокую встречаемость смолевки ползучей и сурепки сжатой, соответственно 65,8 и 57,3% . Относительно высока встречаемость лапчатки вильчатой из растений семейства Розоцветные (4,8%).
В эколого-географическом отношении три ведущих по встречаемости вида сорняков (от 9,7 у пырея ползучего до 65,8% у смолевки ползучей) относятся к мезоксерофитным и мезофитным видам широкого евразиатского распространения; генетически они относятся к бореальным видам и тяготеют к лесной и лесостепной флоре. Из этой группы лишь стержнекорневой монокарпик - сурепка сжатая - относится к типичным сорно-залежным видам, остальные два (смолевка и пырей ползучий) относятся к стержнекорневым и длиннокорневищным многолетникам и по классификации залежных растений (Дубровский и др., 2005) они являются переходными, т.е. свойственны как естественным лугово-степным, так и залежным фитоценозам. Интересную группу слагает второй блок видов в залежной флоре посевов проса, это виды с встречаемостью от 2,4 (полынь однолетняя) до 3,6-4,8% (конопля сорная и лапчатка вильчатая). Они в посевах выделяются в виде локальных куртин, формирующих своеобразную мозаичность агроценозов. Самые низкие показатели встречаемости (1,2%) имеют полынь метельчатая, котовник сибирский и люцерна серповидная. В целом в составе сорной флоры просовых посевов доминируют виды полыней и лапчаток, остальные представлены моновидовыми родами, хотя их сорно-ценотическая роль может быть значительной, как в случае с сурепкой и вьюнком полевым.
3.2. Особенности разнообразия просовых сообществ и эколого-ценотический анализ эталонного полынно-сурепково-просового агроценоза. В ходе маршрутных исследований нами были описаны сообщества на посевах проса. При этом характеристика конкретных агроценозов проводилась в разные фазы вегетации P. miliaceum L., с целью более полного учета состава сопутствующих просу сорняков.
На основе анализа уже 10 геоботанических описаний наблюдается видовая насыщенность в ценофлоре сообществ просовых агроценозов. Классификация растительности просовой формации с выделением ассоциаций построена на эколого-фитоценотической основе (Александрова, 1969). Анализ геоботанических описаний 13 пробных площадок выявил три группы однотипных по доминирующим видам и по структуре сообществ, группы описаний, которые можно трактовать в ранге ассоциаций, в их числе:
- Ас. Сурепково-просовая (Barbarea stricta + Panicum miliaceum)
- Ас. Полынно-просовая (Artemisia annua + Artemisia scoparia - Panicum miliaceum)
- Ас. Вьюнково-смалевково-просовая (Convolvulus arvense + Silene repens - Panicum miliaceum)
В работе дается подробная характеристика выделенных ассоциаций.
Для более глубокого определения эколого-ценотической структуры просовых ценозов был заложен ключевой участок (полынно-сурепково-просовый агроценоз) в Чаа-Хольской долине, в центральной части Тувы. Местность, где возделывается просо, относится к долине р. Сай-Суу. Участок террасы площадью 27,0х143,0 м используется под возделывание проса. Почва - лугово-дерновая легкосуглинистая.
Методика. В посеве проса, занятом полынно-сурепково-просовым агроценозом, методом случайной выборки были заложены линейные трансекты длиной 0,5 м, где учитывалась встречаемость каждого вида. Выборка состояла из 87 трансектов. Всего было отмечено 22 вида высших сосудистых растений. Общее количество зарегистрированных особей - 700. Статистическая обработка данных проведена при помощи программы Excel (Statistica 7.0). Для выявления сопряженных связей видов использовался метод корреляционных плеяд П.В. Терентьева (1960).
В обработку включено 11 видов с встречаемостью более 5%. При анализе корреляционных связей видов в сообществе основное внимание было уделено уровням связей сорных растений по отношению к просу. В результате четко определились группы видов с высокой, средней и слабой степенью корреляции, что отражено в матрице корреляционных связей и выявленных плеядах - группах сопряженных видов (рис. 2).
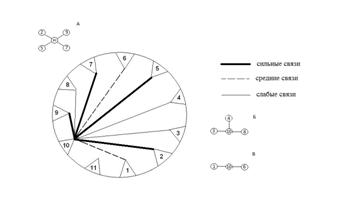
Рис. 2. Схемы плеяд корреляционного цилиндра (1-11 - виды растений, см. табл. 2; А, Б, В - плеяды сопряженных с просом видов). Сильные связи - 0,83 и более; средние связи - 0,71-0,83; слабые связи - 0,41-0,71 а
Анализ сопряженностей видового состава сообщества по отношению к возделываемой культуре проса выявил три плеяды - группы видов с различными уровнями корреляций (рис. 2). Первая плеяда состоит из четырех видов - Elytrigia repens, Silene repens, Convolvulus arvensis, Barbarea stricta; вторая плеяда из трех видов - Medicago falcata, Artemisia annua, Artemisia scoparia; третья плеяда из двух видов - Cannabis sativa, Potentilla bifurca.
В целом небольшое число видов (4 вида из 22 зарегистрированных) в первой плеяде с высокими связями с просом подчеркивает очень слабые ценотические связи в агроценозе. Анализ биоэкологических особенностей видов (табл. 2), входящих в плеяды, позволил дать следующую характеристику каждой плеяде.
Таблица 2
Группы сопряженных видов-сорняков (А, Б, В - плеяды с разными уровнями связей) и некоторые биоэкологические их особенности
Плеяды |
Вид и номер |
Жизнен-ная форма |
Тип корневой системы |
Способ размноже-ния |
Экологичес-кая группа |
Тип ареала |
А |
Пырейползучий (Elytrigia repens) (9) Смолевкаползучая (Silene repens) (7) Вьюнокполевой (Convolvulus arvensis)(5) Сурепкаприжатая (Barbarea stricta) (2) |
Трав. много-летник Трав. много-летник Трав. много-летник Одно-летник |
Длиннокорневищный
Короткокорневищный Стержне-кистекор-невой Стержне- вой |
Семенное и вегетатив-ное Семенное и вегетатив-ное Семенное
Семенное |
Ксеромео-фит
Мезоксеро-фит
Ксерофит
Ксеромезо-фит |
Евразийский
Общеазиатс-кий
Голарктичес-кий
Евразийский |
Б |
юцернасерпови-дная(Medicago falcata) (8) Полыньоднолетняя (Artemisia annua) (4) П. метельчатая (Artemisia scoparia) (3) |
Трав. много-летник. Одно-летник
Однод-вулетник |
Стержне-вой
Стержне-вой
Стержне-вой |
Семенное
Семенное
Семенное |
Ксеромезо-фит
Мезоксеро-фит
Ксерофит
|
Евразийский
Евразийский
Евразийский |
В |
Конопля посевная (Cannabis sativa)(1) апчатка вильчатая (Potentillabifurca) (6) |
Однолетник
Трав. многолетник |
Стержне-вой
Коротко-корневищный |
Семенное
Семенное и вегетативное |
Мезоксеро-фит
Ксерофит |
Голарктиче-ский
Евразийский |
1. В группе видов (плеяда - А) с наиболее сильными связями с просом отмечается отбор тех биоморф, экологические особенности которых наиболее близки к возделываемой культуре. Это преимущественно травянистые многолетники ксеромезофитной экологии с обширным евразийским и голарктическим типами ареалов. В плеяде особняком выделяется сурепка - содоминант в сообществе, имеющая наиболее высокие связи с просом (0,95). Участие этого евразийского ксеромезофитного однолетника с мощной энергией семенного возобновления в какой-то мере указывает на вероятные парагенетические отношения в эволюции проса как культурного растения в центральноазиатском секторе ее ареала.
2. В третьей плеяде (Б) выделяются одно- и двулетние полыни Artemisia scoparia и A. annua, уровень связей с просом - 0,58. Эти виды относятся к ксерофитам с широкими евразийскими ареалами и прекрасным семенным возобновлением. Полыни имеют слабые показатели корреляционных связей с просом при довольно высоких значениях связей (0,72 - 0,74) с другими сорными растениями (Convolvulus arvensis, Nepeta sibirica). Это в какой-то мере указывает на универсальность этих сорняков, поскольку этими полынями бывают засорены посевы многих культурных растений (Марков и др., 1964). Присутствие люцерны (травянистый многолетник ксеромезофитной экологии) в составе этой группы вероятно, связано с предыдущей культурой в севообороте, имеет самый низкий показатель взаимной сопряженности (0,44) с просом. Кроме того, слабая связь люцерны с просом является следствием ее экологии, поскольку она отмечаются в местах с особыми экологическими условиями - к поверхностно-каменистым и локально увлажненным участкам пашни.
3. Вторая плеяда (В) - самая малочисленная и слагается двумя видами (Cannabis sativa, Potentilla bifurca) довольно контрастной экологии. С одной стороны, конопля - сорный однолетник мезоксерофитной экологии с голарктическим распространением, с другой - короткокорневищный травянистый многолетник с более узким евразийским ареалом. Уровень связей у обоих видов с просом - 0,73. Они в посевах выделяются в виде локальных куртин, формирующих своеобразную мозаичность агроценозов. Конопля издавна введена в культуру и относится к привычным сорно-залежным растениям, а в отличие от него короткокорневищный травянистый многолетник - лапчатка вильчатая - еще не утратил ценогенетических связей с исходной степной растительностью (Терехина, 2000).
Таким образом, в составе просовых посевов Центральной Тувы выделяется три ведущих по показателям встречаемости вида сорняков (пырей ползучий, смолевка ползучая и сурепка сжатая). Если первые два вида относятся к мезоксерофитным и ксерофитным видам широкого евразийского распространения, то стержнекорневой монокарпик - сурепка сжатая - относится к типично сорно-залежным видам.
ГЛАВА 4. Биоэкологические особенности Panicum miliaceum L и морфобиологическое разнообразие экотипов проса. Просо обыкновенное (PanicummiliaceumL.) за пределами лесостепной зоны возделывается эпизодически, проникая до 53-54 с. ш. В горах культура просо известна на высотах до 3000 м и более над ур. м. (Тянь-Шань, Памир, Тибет и т.д.) (Вульф, 1945).
Просо по экологии мезоксерофит, это теплолюбивое жароустойчивое короткодневное растение с малым вегетационным периодом, что допускает применение поздних сроков посева, отмечает П.М. Жуковский (1950). При этом только первые фазы вегетации проходят замедленно. Засухоустойчивая и светолюбивая природа проса весьма важна для возделывания этого вида в Туве.
4.1. Особенности фенологического развития проса в Центральной Тыве (на примере опыта возделывания чаа-хольской популяции вида). В работе представлены данные по ритмике развития проса за вегетационный период 2009-2010 гг. на посевах семейно-родового фермерского хозяйства с. Булун-Терек Чаа-Хольского кожууна. Местность, где возделывается просо, относится к долине р. Сай-Суу (левый приток р. Чаа-Холь). Участок террасы реки площадью 27,0 х 143,0 м используется под возделывание проса. Почва - аллювиальная лугово-дерновая, легкосуглинистая, поверхностно корковатая.
В посеве использовались семена местной чаа-хольской (булун-терекской разновидности) популяции проса. Исследовался сезонный цикл развития побегов и репродуктивных структур проса. Знания об эколого-биологичес-ких особенностях различных эколого-географических рас, культивируемых в регионе сорта проса, важны для разработки рекомендаций по их районированию, выработке подходов для селекционного улучшения сорта в климатически контрастных районах Тувы.
Материал и методика. Материал был собран летом 2009/10 г. на опытном поле в окрестности с. Булун-Терек, где выращивали сорт Иртышское-10 (Жуланова, 2005), который уже многие десятилетия культивируется в данной местности. Сегодня она представляет собой особую тувинскую разновидность исследованного сорта (чаа-хольская популяция). В ходе исследований было выявлено два экотипа местной популяции - Чочак-тараа и Хоор-тараа., фенологические стадии их развития были рассмотрены в сравнительном плане. Для сравнения приведены данные по феноритмике нагорной Сут-Хольской популяции проса в условиях лабораторного эксперимента - ex-citu.
Для анализа в условиях ex-citu использовали метод 1000 семян. Для этого отсчитывались две пробы чистых семян проса по 500 штук. Далее взвешивались с точностью до граммов на аналитических весах.
До посева семена (30 штук) предварительно замачивались в воде в течение 12 часов при температуре 25-27оС. Это обеспечивает появление быстрых и дружных всходов. Посев проса производился на каштановой легкосуглинистой почве в глубину 3-4 см.
Результаты изучения биоритмов развития по фенологическим фазам исследованных образцов проса представлены в таблице 3.
Через 4-5/6 дней после посева (табл. 3) всходы экотипа-Б появились а22.06 и 08.06 в виде шильца, способного к вытягиванию. Несмотря на малые размеры, семена проса всходят с глубины 10-12 см. Однако температурный фактор влияет на процессы развития всходов. Так, данные наших лабораторных опытов с семенами местной сут-хольской селекции, показали, что оптимальная температура для их роста и развития - это 12-13C при глубине заделки семян на 3-4 см (Ооржак и др., 2011).
аТаблица 3
Фенологические ритмы развития проса в Чаа-Хольской долине Центральной Туве (2009-2010)
Просо |
Дата посева |
Фазы вегетации |
Вегетационный период, дн. |
||||||
Всходы |
Кущение |
Трубкование |
Колошение |
Цветение |
Спелость |
||||
Восковая |
Полная |
||||||||
Panicummiliaceum L. (чаа-хольская популяция, экотип Б Чочак - тараа.) |
16.06.09/ 03.06.10 |
22.06.09/ 08.06.10 |
07.06.09/ 21.06.10 |
16.07.09/ 01.07.10 |
30.07.09/ 12.07.10 |
14.08.09/ - |
30.08.09/ - |
08.09.09/ 06.09.10 |
79/92 |
Panicummiliaceum L. (чаа-хольская популяция, экотип А Хоор - тараа) |
03.06.09/ 12.05.10 |
08.06.09/ 04.06.10 |
01.07.09/ 17.06.10 |
13.07.09/ 3.07. 10 |
22.07.09/ 14.07.10 |
а- / 22.07.10 |
20.08.09/ - |
06.09.09/ 28.08.10 |
92/86 |
Panicummiliaceum L. (сут-хольская популяция, ex-citu) |
27.01.09 /14.06.10 |
30.01.09/ 19.06.10 |
13.02.09/ - |
22.02.09/ - |
06.03.09 / -
|
- |
а- |
14.04.09/ 31.08.10
|
76/74 |
Примечание. Данные за 2009 год приведены в числителе, а в знаменателе - показатели 2010 года. Тире означает отсутствие сведений.
Эта особенность местных популяций проса очень важна для сухостепных районов Тувы, где нередко пересыхает поверхностный горизонт почвы. Однако с появлением второго и третьего листьев рост стебля несколько приостанавливается и из узла кущения, который закладывается обычно у поверхности почвы, начинают отрастать вторичные корни.
Кущение - особая форма ветвления побегов, характеризующегося образованием скученной группы боковых побегов близ основания главного из наземных, приповерхностных и подземных почек. При хорошем увлажнении в фазе кущения образуется уже достаточно мощная корневая система. В нашем случае кущение наступило через 13 дней после появления массовых всходов и продолжался в течение 11 дней.
По данным М.П. Модестова (вероятно, в условиях полевого опыта), кущение начинается через 15-20 дней после появления всходов и продолжается 10-12 дней. В нашем опыте всходы экотипа-А Хоор-тара на 9-10 дней запаздывали от особей экотипа-Б Чочак-тараа (табл. 3), но, однако, к фазе колошения вступили одновременно, т.е. 12 июля. Это указывает о высоком адаптивном потенциале данного экотипа, который еще не культивируется широко в регионе. В этой фазе вегетации происходит зачаточное формирование стеблей, интенсивное развитие корневой системы (рис. 3). Во время кущения наряду с нарастанием вегетативных органов развиваются и конусы роста, из которых позже образуются метелки.
Кущение при недостатке тепла может растягиваться до 35-40 дней (Якименко, 1975). Наиболее энергично просо кустится при температуре 18-20C и достаточной влажности почвы (60-80% НВ). В целом при дефиците освещенности, тепла и влаги фаза кущения растягивается и в результате образуется много подседа, т.е. развитие побегов без закладки цветковых почек.
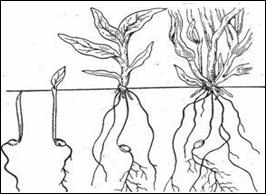
 Посевы проса легко угнетаются сорняками в связи с медленным ростом, особенно в первый период после появления всходов. Именно на данных стадиях заметны синузии сорняков, их яркий аспект во время цветения. В исследованном нами просовом агроценозе ярко-желтый оттенок давала Barbareastricta Andrz. С начала фазы кущения, когда уже достаточно развита мочковатая корневая система проса, рост растений резко ускоряется и сорняки заметно подавляются.
Посевы проса легко угнетаются сорняками в связи с медленным ростом, особенно в первый период после появления всходов. Именно на данных стадиях заметны синузии сорняков, их яркий аспект во время цветения. В исследованном нами просовом агроценозе ярко-желтый оттенок давала Barbareastricta Andrz. С начала фазы кущения, когда уже достаточно развита мочковатая корневая система проса, рост растений резко ускоряется и сорняки заметно подавляются.
При развитии на побегах растений 6-7 листьев они переходят в фазу выхода в трубку. Это связано с усиленным ростом стебля, при котором интенсивно нарастает надземная масса растений и развивается корневая система. В фазу выхода в трубку особи экотипа-Б вступили 1 июля, т.е. на 2 дня раньше, чем экотип-А в 2010 г.
По нашим наблюдениям, в этой фазе увеличиваются размеры как растения, так и конусов нарастания, на нижней части которых появляются бугорки (начинается закладка цветковых почек в соцветии). Образуются веточки-метелки. За 9-10 дней у местной популяции проса зачаточная метелка приобрела форму плотной кисти, затем началась дифференциация колосков и удлинились веточки (на 4-й день). В колосках сформировались чешуйки, цветочные пленки и органы цветка. Интересно отметить, что особи местной популяции проса экотипа-A эту фазу на 2 дня проходит быстрее и, по сути, выравнивается с экотипом-Б (табл. 3).
Выметывание (или колошение) означает процесс выхода соцветия из влагалища верхнего листа. Колошение проса в условиях лабораторного опыта длится от 12 до 18 дней. Цветы в колосках начинают зацветать через 2-4 дня. Как показали наблюдения, продолжительность полного выметывания одной метелки проса составляла от 8 до 14 дней. Интересно отметить, что метелки, заканчивавшие полное выметывание за более короткий срок, отличались большей продуктивностью. К сожалению, в полевом опыте нам не удалось зафиксировать время вступления в фазу начала созревания семян - молочной спелости.
Таблица 4
Климатические показатели за вегетационный период лета 2009-2010 гг.
по данным гидрометеослужбы РТ (по Улуг-Хемскому району)
|
Годы наблюдения |
май |
июнь |
июль |
ав- густ |
сентябрь |
абсол. макс. за период май-сентябрь |
Температура воздуха(t0 C) |
2009 |
12,1 |
18,2 |
28,4 |
29,2 |
11,6 |
28,4 (июль) |
2010 |
11,7 |
18,6 |
22,9 |
28,8 |
10,8 |
28,8 (август) |
|
Температура почвы (t0 C) |
2009 |
52 |
50 |
60 |
60 |
34 |
60 (июль-август) |
2010 |
51 |
61 |
56 |
47 |
44 |
61 (июнь) |
|
Осадки(мм., в месяц) |
2009 |
29 |
94 |
79 |
145 |
44 |
145 (август) |
2010 |
31 |
66 |
80 |
173 |
49 |
173 (август) |
Цветение проса наступило через две недели со дня начала выметывания у экотипа-Б в 2009 г., а в 2010 г. данную фазу особи экотипа-А проходили в июле и ускоренном ритме на 7 дней раньше (табл. 3). Вероятно, это связано с менее благоприятными метеоусловиями середины лета этого года - несколько прохладного по показателям теплообеспеченности как воздуха, так и почв (табл. 4). В целом конец мая и июнь 2010 г. были заметно засушливыми, и это отразилось в ритмике развития - запаздывание в начале вегетации и в дальнейшем ее ускорение, особенно к фазе колошения и цветения (Сводка ГидрометеоцентраЕ, 2011). Август был благоприятным по осадкам и теплообеспеченности (табл. 4). С наступлением цветения у проса приостанавливается рост стебля и развитие корневой системы. Цветение продолжалось 10-12 дней. Первыми зацветали верхние и наружные боковые колоски, затем нижние колоски у основания метелки. Цветение наблюдается в основном утром, между 10-11 часами.
Зерна (семянки) начали созревать, вступая в фазу восковой спелости, на 48-49-й день, и продолжалось дальнейшее созревание зерновок, вплоть до полной спелости в течение 2 недель (15-16 дней), до 6 сентября. При молочной спелости зерна имеют зеленый цвет, при придавливании зерна появляется студенистое белесое образование. Зерна начинали созревать с верхней части метелки, потом в середине и в конце созревали цветки, расположенные в нижней части соцветия.
аВ целом при позднем посеве вегетационный период проса сокращается. Обычно полный вегетационный период у исследованной Чаа-Хольской популяции варьирует в пределах 79-92 дней, а по литературным источникам, он равен 80-90 дням с колебаниями 60-120 дней.
4.2. Некоторые биоморфологические особенности экотипов чаа-хольской популяции проса (анатомо-морфологические особенности листьев, морфология вегетативных и генеративных органов, степень всхожести семян). Сегодня просо в Тыве составляет парадоксальный генетический материал, сочетающий в себе признаки культурных сортов наряду с аборигенными - отборными популяциями местной народной селекции. Этот разнородный по происхождению семенной материал служит основой современной культуры проса в Тыве и поддерживается в небольших семейных фермерских хозяйствах.
Предпринятый нами анализ посевов на примере хозяйств Чаа-Хольской долины Центральной Тувы с позиций выявления особенностей биоморф особей и в последующем семян возделываемой культуры проса обнаружил проявление в них признаков двух экотипов. На уровне биоморф четко выделяются, с одной стороны, более крупные особи c сравнительно толстым и темноватым по оттенку стеблем с достаточно мощной энергией кущения, тремя-четырьмя генеративными побегами (экотип-А), с другой - более тонким и белесо-зеленоватым стеблем с одним-двумя плодущими побегами со средними показателями морфометрии особей (экотип-Б). Семенной материал показал также отличия, где особи первой группы имели сравнительно крупные зерновки, более темновато-коричневые по цвету с блестящими цветочными пленками. По сути, их целесообразно рассматривать в ранге двух экотипов внутри высеваемой популяции проса.
В подтверждение этого дадим некоторые количественные показатели по числу особей и зерновок на учетных площадках площадью в 1 м2. По данным на 07.10.09 г., число особей колебалось на 1 м2 (из 5 повторностей), соответственно: 136; 128; 170; 202; 210. Вес зерновок (в граммах) с этих же площадок изменялся соответственно: 50,65; 49,97; 55,72; 23,01; 54,63.
Кроме этого, было выявлено среднее количество семян с одной особи. В результате получились следующие показатели по их изменчивости: 235; 208; 184; 249; 331. Так, из полученных материалов видно, что на второй площадке (128 особей на 1 м2) несмотря на относительно меньшее число особей, получен высокий выход зерновок (49,97 г на 1 м2). Однако количество зерновок с одной особи в среднем несколько меньше (208), чем у особей экотипов второй группы, что, вероятно, компенсируется более крупными зерновками.
Чаа-Хольские экотипы проса имеют особенности и в анатомической структуре листа. Для анализа брали листья, достигшие полного развития в фазе колошения, и основное внимание уделяли строению мезофилла и эпидермиса как наиболее пластичным признакам в структуре листа.
аМетодика. Поперечные срезы делали бритвой от руки - временные препараты. Размер клеток определяли по методу Т.Н. Годнева и Г.А. Липской и выражали в мкм. Измерения и фотографии проводили на электронных микроскопах ТМ-1000 Tablatop Microscope Hitachi и Axiostar plus (программа Axiovision 4.8).
Просо как растение с типом фотосинтеза С4 в структуре листа имеет некоторые особенности (Буинова и др., 2002). Мезофилл и обкладка образуют два концентрических слоя вокруг проводящих пучков, хорошо видимых на поперечных срезах (рис. 4, 5). Лист проса ксероморфный, амфистоматический, с толстой листовой пластинкой (180,55 мкм). Мезофилл кранцизопалисадный: представлен двумя рядами клеток ткани, радиально расположенных вокруг проводящих пучков, без межклетников. Адаксиальные (20,8 мкм) и абаксиальные (27 мкм) клетки эпидермиса крупные, с нетолстой наружной стенкой, в поперечном сечении овальные. Проводящая система развита хорошо. Центральный проводящий пучок выступает на нижней стороне листа. Значительный объем в толще листовой пластинки имеют пузыревидные (моторные) клетки, несущие функцию водозапасающей ткани и регулирующие свертывание и развертывание листовой пластинки. Моторные клетки значительно выделяются по величине и форме у экотипа-А (рис. 5Б). Эти же клетки у более ксерофитного экотипа характеризуются мелкими размерами и однообразно прямоугольными формами (рис. 4Б).
Листовая пластинка с нижней стороны покрыта редкими волосками, при этом их длина заметно различается. Так, у ксероморфного экотипа-Б в отличие от мезоморфного типа (495 мкм) они достигают 630 мкм. Данный признак - сугубо адаптивный, поскольку в условиях большей инсоляции и сухости экотопов от интенсивной транспирации защищает характер опушения листьев. Кроме этого, более засухоустойчивость особей экотипа-А проявляется в мелкоклеточности и большей извилистости стенок клеток эпидермиса листа (рис. 4А). Это в целом характерно для растений ксерофитов.
К особенностям листа проса следует отнести, в общем, мелкие размеры устьиц в сравнении с другими злаками. Размеры устьиц варьируют от 28,3 до 29,5 мкм на верхнем эпидермисе, в пределах 26,7-27,8 мкм - нижнем. Однако у экотипа-Б в сравнении со второй наблюдается незначительное превышение на 1,2 мкм на адаксиальной стороне и на 1,02 - абаксиальной. На обеих сторонах листа двух экотипов просо устьица немногочисленны и расположены глубже эпидермального слоя.
Аа аа ааБ
 аа
аа 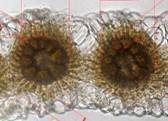
Рис.4. Нижний эпидермис (А) и поперечный (Б) срез листа экотипа-Б (увел.500)
а Б аа а а
 аа
аа 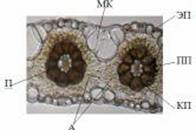 а
а
Рис.5. Нижний эпидермис (А) и поперечный (Б) срез листа экотипа-А (увел. в 500 раз)
ЭП - эпидермис; А Ц ассимиляционная палисадная ткань; МК - моторные клетки; П - паренхимная обкладка; КП - ксилемная паренхима; ПП - проводящие пучки
Выявленные экотипы А и Б в составе местной Чаа-Хольской популяции просо имели некоторые различия по показателям энергии прорастания и по всхожести (табл. 5).
Методика. Энергию прорастания и всхожесть семян вяза определялась согласно ГОСТ 13056.06 - 97 (1998). Проращивание семян просо производилась на чашках Петри.
Для работы с каждой выборки семян двух экотипов отбиралось по 100 семян, в трехкратной повторности. Извлечение проросших семян проводилось в течение всего периода проращивания, количество проросших семян на 5 день определяло энергию прорастания семян, на 10 день - всхожесть.
Так, наиболее высокие темпы прорастания и доля взошедших семян обнаружили зерновки экотипа-Б, при энергии прорастания 97,3% всхожесть была на уровне 98%. Несколько уступает по данным показателям экотип-А. Значительно, более низкие показатели по всхожести и энергии прорастания выявлены у зерновок Сут-Хольской популяции, вероятно более адаптированной к экстремальным условиям Алашского плато. Вероятно, сравнительно низкие показатели всхожести семян компенсируются большой адаптивностью особей к суровым климатическим и эдафическим условиям среднегорий, которые выражаются в частности, в их скороспелости; у них наиболее короткий вегетационный период (табл. 3).
Таблица 5
Энергия прорастания и всхожесть семян проса (в %)
|
Экотипы |
Энергия прорастания |
Всхожесть |
1 |
Чаа-Хольская популяция, экотип-А Хоор-тараа) |
84,0% |
90,5% |
2 |
Чаа-Хольская популяция, экотип-Б Чочак-тараа |
97,3% |
98,0% |
3 |
Сут-Хольская популяция |
70,6% |
77,0% |
Дальнейший анализ разнообразия выявленных экотипов просо показали следующие тренды изменчивости ряда морфобиологических признаков вегетативной и генеративной сферы растения. По данным статистического анализа была построена таблицы 6 по морфометрическим характеристикам двух экотипов просо.
Методика. Исследования проводились 07 июля 2011 г. на просовом агроценозе в урочище Аймырлыг (Чаа-Хольская котловина, терраса р. Сай-Суу). Морфометрические измерения вегетативных и генеративных органов просо проводили на модели 20 случайно выбранных особях по каждому из выявленных экотипов. В качестве показателей вегетативных органов были взяты: H - высота центрального побега особей, T - толщина стебля в средней части основного побега, L - длина листа, D - ширина листа; из генеративных: N - длина цветоножки, Х - длина бутона, У - ширина бутона.
В результате исследования достаточно четко проявились отличия по ряду метрических показателей у исследованных экотипов. Так, экотип-А от экотипа-Б отличается более опушенными и темноватыми побегами, стебли толще почти на 0,1 см, высота основного стебля на 3,1 см выше и в такой же степени длина и ширина листьев превышают показатели экотип-Б (табл. 6). Однако эти признаки наиболее вариабельные, особенно по толщине стебля и ширине листьев. Относительно менее изменчивы показатели по высоте растения. Среди генеративных признаков более стабильными оказались размеры бутонов, особенно их длина. Вся выборка измеренных бутонов была равна 2,0 мм. Незначительное варьирование было обнаружено по ширине бутонов. Высокая степень изменчивости наблюдается по такому показателю, как длина цветоножки, особенно значительное варьирование было обнаружено у особей экотипа-Б.
Полученные сведения по изменчивости морфологических признаков у особей исследованной популяции проса подтвердили, с одной стороны, наличие в просовом агроценозе двух экотипов, с другой - монолитность популяции проса в рамках единого и стабильного вида - Panicummiliaceum L. (Монгуш, Намзалов, 2010).
Признаки цветка как наиболее консервативной и устойчивой части в систематике растений и на этот раз четко проявились. Изменчивость более всего коснулась признаков вегетативных органов растения. Однако выявившиеся экотипы внутри чаа-хольской популяции проса в дальнейшем могут получить развитие и закрепиться в генотипе, особенно перспективным представляется экотип-А.
Таблица 6
Сравнительные морфометрические характеристики просо экотипа-А
c толстым и темноватым стеблем и экотипа-Б с тонким
и светлым стеблем (в см)
аМорфометрия экотипов Морфологические признаки |
Х |
Х min |
Х max |
t |
Cv, % |
Высота растения, H |
42,7/39,4 |
36/31 |
50/47 |
5,88/4,35 |
а13,79/11, 04 |
Толщина стебля, T |
0,37/0,28 |
0,3/0,2 |
0,5/0,3 |
0,06/0,28 |
18,24/15,06 |
Длина листа, L |
23,3/21,5 |
19/17 |
29/26 |
а3,59/2,67 |
15,41/12,45 |
Ширина листа, D |
1,21/0,95 |
1,0/0,6 |
1,6/1,1 |
0,21/0,14 |
18,04/15,09 |
Длина цветоножки, N |
1,25/1,07 |
1,0/0,7 |
1,4/1,4 |
0,15/0,21 |
12,07/19,86 |
Длина бутона, Х |
0,2/0,2 |
0,2/0,2 |
0,2/0,2 |
2,9/2,92 |
0,00/0,00 |
Ширина бутона, У |
0,49/0,36 |
0,4/0,3 |
0,5/0,4 |
а0,03/0,04 |
6,45/13,16 |
аПримечание. X - средние значения, Х min - X max - лимиты признака; t - стандартное отклонение; Cv - коэффициент вариации. В числителе данные экотипа А, в знаменателе - экотипа Б
а
В дальнейшем в целях сохранения генофонда проса и его разнообразия в культурной флоре России и Центральной Азии, необходимы более детальные исследования местных популяций возделываемой культуры.
ГЛАВА 5. Особенности традиционного возделывания и использования проса. Орошение пашни проводилось один раз до посева и от одного до пяти раз за вегетационный период, после появления всходов. Длительность увлажнения участка зависела от почвенных условий. Так, сухие песчаные почвы размачивали один-два дня, глинистые - до пяти-шести суток. После орошения участка, когда земля несколько просыхала, но была еще сырой (обычно через два-три дня), производили посев вручную - взяв горсть, равномерно разбрасывали семена. Некоторые тувинцы предпочитали высевать семена, предварительно вымоченные, другие - сеяли сухие семена. После того как весь участок был засеян, его заделывали андазыном (сохой), присыпая семена землей. Глубина такой поверхностной вспашки составляла 8-12 см, ширина 10-12 см (Вайнштейн, 1972).
Для пахоты тувинские крестьяне употребляли андазын - деревянное приспособление, согнутое под почти прямым углом. На один его конец был посажен металлический сошник, а на другом укреплялся брусок, служивший рукоятью. Таким андазыном землю можно было вспахивать неглубоко, что было оправданно в местных условиях. Боронили колючими и засохшими кустами караганы (чаще всего использовали Caraganabungeiи C. spinosa), привязав его к поперечной балке в один ряд. В качестве рабочих орудий служили лопаты хуурек, мотыги кускунхай и землекопалки озук, а расчистку вели деревянным гребком хетпе (Потапов, 1969).
После краткого исторического экскурса дадим характеристику уборки урожая проса 2010 г. в семейно-родовом хозяйстве Б.М. Шаравии. На поле отмерили участок размером 5х5 м. Жатву провели ручным серпом и ножами. При этом срезается только метелка с захватом лишь небольшой верхушечной части цветоносного побега (примерно 5,0 см.). Метод такой уборки называется молдуруктаар. В результате с деляны площадью 25 м2 был собран первоначальный урожай проса с метелками. Объем собранного урожая заполнил 1 стандартный мешок. На выполнение данного этапа работы ушло 2,5 часа.
Следующий этап работы - освобождение зерновок от колосков в метелках. Для того чтобы отделить зерна от колосковых чешуй, цветоножек, листьев и частей побегов, совершается процедура сильного механического воздействия на наполненный зерном и крепко связанный мешок. Для этого ритмично бьют по мешку палкой особой конструкции. Такой инструмент имеет ручку, которая посредством крепкой веревки соединяется с ровной, как черенок лопаты, рабочей частью инструмента. Данная процедура в процессе сбора урожая называется бажын дужурер (тув.). На данный вид работы примерно затрачивается 40 мин.
Третий этап - этап очистки от колосковых и цветковых чешуй (шелухи). Для этого вываливали все содержимое мешка на полог и начинали развеивать пустым мешком, как большим веером, пока зерна не отделялись от шелухи. Данная процедура очистки зерен называется - лчелбиир.
В целях достижения максимальной очистки зерен дополнительно провеивали семянки, используя какую-либо широкую посуду (миску), которую наполняли наполовину и высоко поднимали вверх над пологом, медленно высыпали так, чтобы шелуха отвеивалась по направлению ветра. После 1,5 часа такой скрупулезной работы получали чистые зерна проса весом 9,6 кг. Это конечный очищенный продукт от мешка сырого первичного урожая.
Тувинцы заготавливали на зиму 2-3 шан зерна (1 шан = 2 барба; 1 барба = 3 мешка или в пересчете на чистый урожай она составляет 150-170 кг). Таким образом, в среднем получали урожай от 300 до 700 кг проса. В более благоприятные годы суммарный урожай в этих небольших семейно-фермерских хозяйствах достигал одной тонны.
Растительная пища занимала и продолжает занимать большое место в питании тувинцев. Просо - главная их зерновая культура, которая выращивалась на протяжении многих веков. Поджаренное просо и талкан, как угощение для гостей является традиционным и почетным. Чинге-тараа - продукт из проса, является основной составляющей для приготовления традиционных блюд. Приготовление чинге-тараа включает несколько этапов:
- Необрушенное просо замачивали до разбухания, но при этом следили, чтобы оболочка зерна не трескалась. Для убыстрения процесса варили в кипящей воде.
- Затем воду сливали, а вымоченное просо жарили в сухом котле (по порциям), постоянно помешивая кусочком войлока или плотной ткани, намотанной клубком на конце жерди (1,3 м).
- Зерно высыпали в деревянное корыто - деспи - для остужения.
- Небольшими порциями зерна просо (2 пиалы) обрушивают в деревянной ступе согааш при помощи песта бала до первого толчения (кара дук). После первого отвеивания оно еще засорено шелухой, поэтому подвергают толчению в ступе второй раз.
- Высыпают в корыто и подвеивают второй раз, после чего отделяется зерно ак-дук (белый волосок-досл.) и при этом цвет зерна светлый, оболочка тонкая.
- Чистый арыг-тараа получают после третьего обрушивания и подвеивания на корыте.
Следует отметить, что общеизвестный продукт проса в виде каши не столь популярен в Тыве. Чаще из проса делают блюдо оремелиг тараа, приготовленное из полуфабриката чинге-тараа, которое принимается в пищу со сливками или в сочетании с пенкой (типа мюсли). Это очень вкусное и питательное блюдо. Возможно, что мюсли - любимое в Европе блюдо, впервые было приготовлено предками современных тувинцев.
Для выявления ценных пищевых качеств чинге-тараа были проведены исследования химического состава блюд, приготовленных из проса разного происхождения: 1 - чинге-тараа из урожая проса чаа-хольской популяции;
2 - чинге-тараа из проса сорта саратовский. Образцы проса подвергались стандартной обработке по традиционной технологии, начиная от замачивания в воде, прожаривания в котле при одинаковых температурах и времени, толчения в ступе в несколько этапов. Затем у образцов готовых блюд исследовали химический состав (табл. 7)
Таблица 7
Химический состав образцов блюда чинге-тараа
из проса различного происхождения
Образцы проса |
Протеин % |
Содержание воздушно-сухого вещества, в кг |
||
Кальций, г |
Фосфор, г |
Калий, г |
||
1. Чаа-хольская популяция |
11,25 |
1,0 |
2,8 |
3,3 |
а2. Просо сорта саратовский |
9,44 |
0,6 |
1,8 |
2,4 |
Примечание. Химические анализы выполнены в лаборатории ФГБУ Государственная станция агрохимической службы УТувинскаяФ.
аИз таблицы 7 видно, что зерна проса, готовые к употреблению, различаются по химическому составу. Так, содержание протеина в чинге-тараа, приготовленном из чаа-хольского проса, - 11,25%. Содержание кальция и калия также преобладает у чаа-хольского образца. Следует отметить, что зерна проса сорта саратовский во время приготовления продукта по традиционной технологии разрушились и обрели вид муки. Именно чаа-хольская популяция проса наиболее приспособлена к приготовлению традиционных блюд тувинской кухни. Однако в Тыве возделывают просо на очень небольших площадях и лишь в 5 кожуунах (Сут-Хольском, Барун- Хемчикском, Дзун-Хемчикском, Улуг-Хемском, Чаа-Хольском). Необходимо возрождение этой ценной и традиционной культуры в Туве и для этого важно изучить разнообразие генофонда и экологию местных популяций вида для дальнейшей селекции перспективных форм.
ВЫВОДЫ
1. Флора сорняков в посевах проса в Центральной Тыве насчитывает 41 вид сосудистых растений (31 род и 19 семейств), где основу составляют виды монокарпики с обширными голарктическими и евразийскими ареалами, ксеромезофитной и мезоксерофитной экологии, что в целом свойственно адвентивной флоре агрофитосистем.
2. Анализ разнообразия просовых сообществ с использованием эколого-фитоценотического подхода позволил выявить три ассоциации: сурепково-просовая, полынно-просовая и смалевково-вьюнково-просовая, которые отличаются по комплексу дифференциальных и сопутствующих видов, а также по структуре. Выделенные синтаксоны приурочены к определенным элементам горно-долинного рельефа, а также по видовому составу связаны с ценозами, предшествовавшими современным посевам проса.
3. Эколого-ценотическая структура просовых сообществ характеризуется простотой сложения, с формированием маловидовых группировок - плеяд видов. Среди видов, положительно сопряженных с просом, выделяется ксеромезофитный евразийский монокарпик Сурепка сжатая, в сочетании со смолевкой ползучей - короткокорневищным травянистым многолетником азиатского распространения, что, вероятно, связано с их общей адаптивной экологией.
4. В ходе эколого-биологических исследований проса чаа-хольской популяции было отмечено развитие двух экотипов, с характерными морфологическими признаками в структуре вегетативных и генеративных органов, а также анатомическим строением листьев. Кроме этого, выявленные экотипы обнаружили особенности в фенологии развития, а также по морфологии зерновок и по урожайности, которые будут полезными в селекционной работе с местными популяциями проса.
5. В результате анализа опыта возделывания проса в Тыве выявлено не только разнообразие форм в местных популяциях вида, но и удалось раскрыть особенности этнических традиций возделывания и использования проса, уборки и хранения урожая, способов приготовления оригинальных блюд национальной кухни, что является ценным вкладом в познание одного из древнейших пищевых растений человечества.
Список опубликованных РАБОТ
- Намзалов Б.Б. Просо в Тыве: наблюдения и размышления / Б.Б. Намзалов, .К. Монгуш, Н.Г. Дубровский // Проблемы ботаники Южной Сибири
и Монголии: материалы VIII Междунар. науч.-практ. конф. (г. Барнаул, 19-22 октября 2009 г.). - Барнаул, 2009. - С. 312-315.
- Монгуш Л.К. Из истории развития земледелия и возделывания проса
в Туве / Л.К. Монгуш // Актуальные проблемы исследования этноэкологических и этнокультурных традиций народов Саяно-Алтая: материалы I Межрегион. науч.-практ. конф. молодых ученых, аспирантов и студентов (г. Кызыл, 30 июня - 3 июля 2009 г.). - Кызыл: Изд-во ТывГУ, 2009. - С. 28-31.
- Монгуш Л.К.Особенности агроценозов проса посевного в Центральной Тыве и традиционные технологии приготовления продуктов питания /Л.К. Монгуш, Б.Б. Намзалов // Традиционное хозяйство в системе этноса: материалы IX Санкт-Петербургских этнографических чтений (г. Санкт-Петербург, 6-8 декабря 2010 г.). - СПб., 2010. - С.119-127.
- Монгуш Л.К. О внутрипопуляционной изменчивости проса в Тыве /
Л.К. Монгуш, Б.Б. Намзалов // Проблемы экологии: чтения памяти проф.
М.М. Кожова: тез. докл. междунар. науч. конф (г. Иркутск, 20-25 сентября 2010 г.). - Иркутск: Изд-во Иркут гос. ун-та, 2010. - С. 154.
- Монгуш Л.К. Сорняки в посевах проса в Тыве: анализ биоморфологического и эколого-географического разнообразия / Л.К. Монгуш, Б.Б. Намзалов // Вестник БГУ. Сер. Биология. География. - Улан-Удэ, 2010. - Вып. 4. - С. 140-144 (реценз.).
- Ооржак А.В. Особенности фенологического развития проса (сорт тувинское местное): опыт исследования exsitu / А.В. Ооржак, .К. Монгуш,
Л.В. Будажапов // Вестник БГУ. Сер. Биология. География. - 2011. - Вып. 4. -
С. 85-87. (реценз.)
- Монгуш Л.К. Об особенностях возделывания проса и традициях приготовления продуктов из него в Туве / Л.К. Монгуш // Современные проблемы этноэкологии и традиционного природопользования: материалы всерос. науч.-практ. конф. (г. Улан-Удэ, 6-7 декабря 2010 г.). - Улан-Удэ: Изд-во Бурятского госуниверситета, 2011. - С. 65-70.
- Монгуш Л.К. О некоторых особенностях структуры полынно-сурепково-просового агроценоза в Центральной Тыве / Л.К. Монгуш, Б.Б. Намзалов // Сорные растения в изменяющемся мире: актуальные вопросы изучения разнообразия, происхождения, эволюции: материалы Первой междунар. науч. конф. (г. Санкт-Петербург, 6-8 декабря 2011 г.). - СПб.: Изд-во Всероссийского НИИ растениеводства им. Н.И. Вавилова, 2011. - С. 235-241.
- Монгуш Л.К. О некоторых этноэкологических аспектах традиционного возделывания проса в Центральной Тыве / Л.К. Монгуш, А. Бурбужап // Агрономическая наука - взгляд в будущее: материалы науч.-практ. конф. - Улан-Удэ: Изд-во БГСХА, 2011. - С. 32-35.
Подписано в печать 20.04.12. Формат 60?84 1/16.
Усл. печ. л. 1,5. Тираж 100 экз. Заказ 87.
Издательство Бурятского государственного университета
670000, г. Улан-Удэ, ул. Смолина, 24 а
riobsu@gmail.com
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]