
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Пластичность нигростриатной дофаминергической системы при моделировании паркинсонизма у мышей
Автореферат кандидатской диссертации
На правах рукописи
Козина Елена Александровна
ПЛАСТИЧНОСТЬ НИГРОСТРИАТНОИ ДОФАМИНЕРГИЧЕСКОИ СИСТЕМЫ ПРИ МОДЕЛИРОВАНИИ ПАРКИНСОНИЗМА У МЫШЕЙ
03.03.01. - физиология
Автореферат
диссертации на соискание ученой степени кандидата биологических наук
Москва-2012
Работа выполнена в лаборатории гормональных регуляций Федерального государственного бюджетного учреждения науки Института биологии развития им. Н.К. Кольцова Российской академии наук
Научный руководитель:
доктор биологических наук, профессор, академик РАН Угрюмов Михаил Вениаминович
Официальные оппоненты:
Раевский Владимир Вячеславович - доктор биологических наук, профессор, Федеральное государственное бюджетное учреждение науки Институт высшей нервной деятельности и нейрофизиологии Российской академии наук, руководитель лаборатории нейроонтогенеза
Ковалев Георгий Иванович - доктор медицинских наук, профессор, Федеральное государственное бюджетное учреждение Научно-исследовательский институт фармакологии имени В.В. Закусова Российской академии медицинских наук, руководитель лаборатории радиоизотопных методов исследований
Ведущая организация:
Федеральное государственное бюджетное учреждение Московский научно-исследовательский институт психиатрии Министерства здравоохранения и социального развития Российской Федерации
Защита диссертации состоится л 18 апреля 2012 г. в 1400 часов на заседании Диссертационного совета Д 002.238.01 при Федеральном государственном бюджетном учреждении науки Институте биологии развития им. Н.К. Кольцова Российской академии наук по адресу: 119334, Москва, ул. Вавилова, 26
С диссертацией можно ознакомиться в библиотеке Федерального государственного бюджетного учреждения науки Института биологии развития им. Н.К. Кольцова Российской академии наук по адресу: 119334, Москва, ул. Вавилова, 26
Автореферат разослан ла марта 2012 г.
Ученый секретарь Диссертационного совета,
кандидат биологических наука sE.Б. Абрамова
e-mail:а ele0806(g>vandex.raа Q)js^yаа /
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Одним из наиболее распространенных нейромедиаторов в мозге является дофамин. Нарушение метаболизма дофамина при гибели синтезирующих его нейронов у человека приводит к развитию одного из тяжелейших нейродегенеративных заболеваний - болезни Паркинсона (БП) {Bernheimer et al., 1973; Agid, 1991). В основе патогенеза БП лежит деградация дофаминергической нигростриатной системы мозга, обеспечивающей регуляцию двигательных функций. Характерной особенностью БП является длительное бессимптомное течение на протяжении многих лет, что, предположительно, обусловлено включением механизмов пластичности мозга. Появление со временем симптомов БП связывают не только со значительной деградацией нигростриатной системы, но и с практически полным истощением компенсаторных резервов мозга (Agid, 1991).
В силу невозможности изучения эндогенных процессов, протекающих в мозге человека на досимптомной стадии БП, вопрос о развитии нейродегенеративных и компенсаторных процессов на ранних стадиях заболевания до сих пор остается открытым и является актуальной проблемой нейронауки. Важно отметить, что фармакотерапия БП, которая начинает проводиться только после появления двигательных нарушений, со временем становится менее эффективной и приводит к появлению ряда осложнений (Mones et al., 1971; Poewe, 2006). В данной ситуации становится очевидной необходимость разработки доклинической диагностики и превентивного лечения БП. Однако создание подобных технологий невозможно без комплексного изучения механизмов пластичности мозга на ранней досимптомной стадии заболевания. Единственно возможным способом изучения ранних нейродегенеративных и компенсаторных изменений в мозге является экспериментальное моделирование БП на животных, причем основной акцент при моделировании необходимо сделать на длительное развитие нейродегенеративных процессов. В большинстве экспериментальных исследований БП, создаются модели, при которых у животных в течение короткого времени развивается симптомная стадия заболевания. При таких способах моделирования невозможно оценить не только компенсаторные процессы, включающиеся на длительной досимптомной стадии, но и те изменения, которые запускают переход заболевания из одной стадии в другую. Именно эти изменения могут служить в дальнейшем мишенью для поиска лекарственных средств, предотвращающих появление симптомов заболевания.
3
Таким образом, вопрос о разработке экспериментальной модели пролонгированной досимптомнои стадии БП до сих пор остается актуальным, причем особый акцент при моделировании необходимо сделать на медленное и постепенное развитие патологических процессов в нигростриатной дофаминергической системе.
Цель и задачи исследования. Целью данной работы являлась разработка экспериментальнойаа моделиаа пролонгированнойаа воаа времениаа досимптомнои стадии БП, а также изучение механизмов пластичности мозга на досимптомнои стадии различной протяженности (коротко- и длительно-развивающейся). Для достижения цели были поставлены следующие задачи:
- Са помощьюа нейротоксин 1-метил-4-фенил-1,2,3,6-тетрагидропиридин (МФТП) разработать модель пролонгированного развития БП.
- На моделях досимптомнои и симптомной стадий БП в черной субстанции и в стриатуме оценить:
а)а содержание моноаминов и их метаболитов;
б)а количество тел дофаминергических нейронов и терминалей их
аксонов;
в)а содержание мРНК и белка тирозингидроксилазы, а также ее
ферментативную активность.
г)а Содержание мРНК Д2-рецепторов
- Провести анализ иннервации дорзального стриатума при моделировании БП.
- Оценитьаа метаболизмаа МФТПаа ваа дофаминергическойаа нигростриатной системе при его хроническом введении.
Научная новизна. Впервые путем введения нейротоксина МФТП в течение длительного времени смоделировано несколько последовательных фаз хронической досимптомнои стадии БП и ее дальнейший переход в симптомную стадию. Получен ряд важных данных о замедлении скорости развития нейродегенеративного процесса, а именно - замедлении скорости снижения уровня дофамина и дегенерации дофаминергических терминалей в стриатуме, а также активации компенсаторных процессов в нигростриатной системе и за ее пределами при длительном введении МФТП.
Впервые показано, что при моделировании БП с помощью МФТП у животных развиваются адаптационные процессы, происходящие на уровне изменения захвата нейротоксина в дофаминергические нейроны и направленные на выработку устойчивости нейронов к токсическому действию МФТП.
4
Научная и практическая значимость работы. Разработанная модель хронической досимптомной стадии БП в дальнейшем может быть использована для:
а)а поиска периферических эндогенных маркеров досимптомной стадии БП;
б)а разработки превентивной лекарственной терапии, основанной на
снижении чувствительности нейронов нигростриатной системы к
действию экзогенного или эндогенного токсина и направленной на
остановку или замедление дегенерации дофаминергических нейронов.
ичное участие автора. Автором были спланированы и выполнены все эксперименты, описанные в работе, за исключением части экспериментов по анализу моторного поведения животных и высокоэффективной жидкостной хроматографии, которые были проведены совместно с Институтом патологии и общей патофизиологии РАМН и Институтом фармакологии РАМН. Поставленные задачи были решены с применением современных методов физиологии, клеточной и молекулярной биологии. Выводы сформулированы на основе собственных оригинальных данных.
Апробация работы. Материалы диссертации были обсуждены на российских и международных симпозиумах: конференция Механизмы нервных и нейроэндокринных регуляций (Москва, 2008); л2-й Съезд физиологов СНГ (Кишинев, 2008); конференция Нейрохимические механизмы формирования адаптивных и патологических состояний мозга (Санкт-Петербург, 2008); л39th annual meeting Society for Neuroscience (Чикаго, 2009); The Second International Workshop on Image Mining - Theory and Applications (Lisboa, 2009); XXI съезд Физиологического общества им. И.П. Павлова (Калуга, 2010); II Национальный Конгресс по болезни Паркинсона и расстройствам движений (Москва, 2011); л41st annual meeting Society for Neuroscience (Вашингтон, 2011).
Публикации. По теме диссертации опубликовано 5 статей в рецензируемых журналах из списка ВАК, 1 статья в зарубежном рецензируемом журнале, 1 глава в коллективной монографии и 27 тезисов конференций.
Работа поддержана грантами: РФФИ 09-04-12106-офи_м; Российский фонд гуманитарных наук 09-06-00543а; Научные школы 2110.2008.4; Программа Президиума РАН "Фундаментальные науки - медицине"; РФФИ-10-04-93108-НЦНИЛа.
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, описания материалов и методов исследования, результатов,
5
обсуждения, выводов и списка литературы, включающего 342 источника. Работа изложена на 142 страницах машинописного текста, содержит 38 рисунков и 9 таблиц.
СОДЕРЖАНИЕ РАБОТЫ Материалы и методы исследования
Животные, схема эксперимента. В работе использовано 260 мышей-самцов линии C57BL/6 в возрасте 2-2,5 месяца и весом 22-26 г на момент начала эксперимента. Для моделирования хронически развивающегося паркинсонизма животным однократно раз в неделю подкожно вводили МФТП (Sigma, США) в дозе 12 мг/кг в течение 5, 7 и 10 недель (обозначается как 5x12, 7x12, 10x12). В контроле животным, по аналогичной опытным животным схеме, вводили физиологический раствор (0.9% NaCl). В экспериментах по определению активности тирозингидроксилазы (ТГ) опытным и контрольным животным за 30 минут до декапитации внутрибрюшинно вводили ингибитор второго фермента синтеза дофамина (ДА) - декарбоксилазы ароматических L-аминокислот (ДАА) - 3-гидроксибензилгидразин (NSD-1015) (Sigma, США) в концентрации 100 мг/кг (Carlsson, 1973).
Для воспроизведения модели быстроразвивающегося паркинсонизма (лострая модель) использовались следующие схемы введения МФТП: (а) двукратно с 2-х часовым интервалом в дозе 12 мг/кг для моделирования поздней досимптомной стадии (обозначается как 2x12); (б) четырехкратно с 2-х часовым интервалом в дозе 12 мг/кг для моделирования ранней симптомной стадии (обозначается как 4x12).
Оценка поведения. В хронических экспериментах каждые 7 дней (накануне перед следующей инъекцией МФТП) животных тестировали в "открытом поле" (Opto-Varimex-3, "Columbus instruments", США). В течение 3-х минут измеряли пройденный путь, время без движений и число вертикальных стоек {Kryzhanovsky et al. 1997). Дополнительно определяли длину шага (Tillersonet al. 2003). Используемые поведенческие тесты позволяют выявить основные симптомы БП - гипокинезию, атаксию и ригидность (MuralikrishnanandMohanakumar1998; Sedelis et al., 2001).
Взятие материала. В хронических экспериментах сбор материала проводили через неделю после последней инъекции МФТП. Животных из экспериментальных и контрольных групп декапитировали, извлекали мозг и разрезали его по среднесагиттальной плоскости. Из правой половины мозга выделяли черную субстанцию (ЧС) и стриатум {Козина и др., 2010), а затем в образцах измеряли содержание моноаминов с помощью высокоэффективной
6
жидкостной хроматографии с электрохимической детекцией (ВЭЖХ-ЭД). Левую половину мозга фиксировали иммерсией в 4% параформальдегиде, промывали в фосфатно-солевом буфере, инкубировали в 20%-й сахарозе и замораживали в гексане (Lecbiopharm, Россия). Для проведения ПЦР и определения активности ТГ, ЧС и стриатум выделяли из обоих полушарий мозга. В экспериментах с использованием острой модели БП (2x12 и 4x12) сбор материала проводили через 2 недели после введения МФТП.
ВЭЖХ. Определение содержания ДА, метаболитов ДА дигидроксифенилуксусной кислоты (ДОФУК), гомованилиновой кислоты (ГВК), а также норадреналина (НА), серотонина (5-гидрокситриптамин, 5-ГТ) и метаболита серотонина - 5-гидроксииндолуксусной кислоты (5-ГИУК) в ЧС и в стриатуме проводили на обращённо-фазной колонке ReproSil-Pur, ODS-3, 4x100 мм с диаметром пор 3 мкм (Dr. Majsch GMBH, Элсико, Россия) в 0.1М цитратно-фосфатном буфере, содержащем 1.1 мМ октансульфоновую кислоту, 0.1 мМ ЭДТА и 9% ацетонитрил с последующей электрохимической детекцией на стеклоугольном электроде (+0.85 V). В экспериментах по определению активности ТГ в собранных образцах оценивали содержание L-3,4-дигидроксифенилаланина (L-ДОФА), которое является показателем активности данного фермента.
Кроме этого, после десяти недель введений МФТП в опыте и 0.9% NaCl в контроле определяли содержание 1-метил-4-фенил-пиридин иона (МФГГ) в ЧС и в стриатуме. Для этого всем животным на 11-й неделе дополнительно однократно вводили МФТП в дозе 12 мг/кг и через 20 минут после инъекции выделяли ЧС и стриатум. Определение содержания МФП+ проводили при помощи ВЭЖХ-УФ по модифицированной методике, описанной ранее Кавасаки с соавторами (Kawasakiet al., 2007). Измерение проводили на хроматографе Agilent 1100 с дио дно-матричным детектором при длине волны поглощения 295 нм. В состав мобильной фазы входили 50 мМ КН2Р04 и 15% ацетонитрил. Все биохимические данные представлены в виде изменения содержания измеряемых веществ в опыте по отношению к контролю, принятому за 100%.
Стереотаксические инъекции. Для анализа иннервации стриатума при моделировании БП через 2 недели после четырехкратного введения МФТП в дозе 12 мг/кг (4x12) в опыте и 0.9% NaCl в контроле проводили стереотаксические инъекции флуоресцентного ретроградного трейсера в стриатум. Инъекции (0.5 мкл) 10% раствора трейсера (FluoroRuby, Invitrogen, США) проводили в область дорзального стриатума правой половины мозга.
7
Через неделю после инъекции трейсера проводили транскардиальную перфузию сначала 0.02 М фосфатно-солевым буфером в течение 15 минут, а затем охлажденным (до +4С) 4% параформальдегидом на 0.2 М фосфатном буфере в течение 15 мин. После этого мозг постфиксировали погружением в тот же фиксатор, промывали в 0.02 М фосфатно-солевом буфере, инкубировали в 20% сахарозе и замораживали в гексане.
Иммуноцитохимия. Для моно-иммуномечения ТГ на криостате Leica (Leica СМ1850, Германия) приготавливали фронтальные серийные срезы ЧС толщиной 20 мкм и фронтальные срезы стриатума толщиной 12 мкм. В работе использовали кроличьи поликлональные антитела к ТГ (1:2000) (предоставлены проф. Тибо, Франция), вторые биотинилированные антитела (1:200) (Vector Laboratories, США) и авидин-биотиновый комплекс, связанный с пероксидазой (Vector Laboratories, США). Пероксидазу авидин-биотинового комплекса выявляли инкубацией с диаминобензидином (Sigma, США) и Н202.
Для двойного иммунофлуоресцентного мечения ТГ и ДАА в аксонах стриатума на плавающих срезах толщиной 30 мкм использовали овечьи поликлональные антитела к ТГ (1:700) (Chemicon, США), поликлональные антитела к ДАА (1:300) (Abeam, США), вторые антитела, меченные флуоресцеинизотиоционатом (FITC) (1:40) (Sigma, США) и вторые антитела, меченные цианином (СуЗ) (1:500) (Sigma, США).
Микроскопия, анализ изображений. Срезы ЧС после моно-иммуномечения на ТГ исследовали в микроскопе Olympus ВХ51 (Япония), оснащенном цифровой камерой Olympus DP70 (Япония), при увеличении объектива ???. Изображения срезов анализировали с помощью программы AnalySIS5.0. (Olympus, Япония): на каждом срезе обводили область компактной части ЧС, содержащей тела ДА-ергических нейронов, после чего подсчитывали число нейронов, причем только с видимым ядром. Для исключения двойного подсчета нейронов, расположенных на соседних срезах, использовали метод Аберкромби {АЪегсготЫе, 1946).
Для количественного анализа терминалей аксонов срезы стриатума после двойного иммуномечения на ТГ и ДАА исследовали с помощью конфокального микроскопа Leica TCS SPI (Германия). Полученные в конфокальном микроскопе лоптические срезы толщиной 0.2 мкм анализировали с помощью программы ImageJ (NIH, США). Аксональные терминали подсчитывали в дорзальном стриатуме на площади среза 900 мкм2.
Для полуколичественного анализа ТГ в нейронах ЧС на каждом десятом срезе обводили все нейроны в компактной части ЧС. Оптическую плотность
8
нейронов, коррелирующую с концентрацией ТГ, определяли как луровень серого по следующей формуле: оптическая плотность = log (уровень серогоф0на /уровень серогОсигнала) Для полуколичественного анализа ТГ в волокнах стриатума использовалась разработанная совместно с Вычислительным Центром им. А.А. Дородницына РАН в ходе данной работы программа Striatum Image Analysis (Gurevich et. al., 2010).
ПЦР в реальном времени. Содержание мРНК ТГ и Д2-рецепторов в ЧС и в
стриатуме оценивали методом ПЦР в реальном времени. После выделения РНК
из образцов проводили реакцию обратной транскрипции с помощью кита
RevertAidЩ Н Minus First Strand cDNA Synthesis Kit (Fermentas, США), после
чего проводили ПНР с использованием интеркалирующего красителя
S YBRGreen (Maxima S YBR Green Kit, Fermentas) на приборе IQ5 Cycler (Biorad,
США). Для определения содержания мРНК ТГ использовали следующие
праймеры: 5'-CGCTGGATACGAGAGGCATAG-3',а 5'
GTCAGAGGAGCCCGAGGTC-3' (Синтол, Россия). Для определения содержания мРНК Д2-рецепторов использовали следующие праймеры: 5'-GTGGTGACTGGGAGGGATGG-3', 5'-GTGAACAGGCGGAGAATGGATG-3\ При стандартизации реакции определяли содержание мРНК актина (housekeeping gene) по двум последовательностям праймеров: 5'-CCAGAGGCATACAGGGACAGC-3' и 5'-TGGCACCACACCTTCTACAATG-3'. Количественную разницу в экспрессии между опытными и контрольными образцами определяли по формуле: 2"??0(?)
Определение активности транспортера дофамина в срезах мозга ex vivo. После десяти недель введений МФТП для определения активности транспортера ДА использовали метод инкубации срезов ЧС и стриатума с меченым тритием ДА ( Н-ДА), описанный ранее (Хакимова и др., 2011).
Статистическая обработка результатов. Полученные данные обрабатывали статистически с помощью F-теста для определения однородности выборки и t-теста Стьюдента для определения достоверности различий.
Результаты и обсуждение Двигательная активность животных. Через неделю после инъекций МФТП в дозе 12 мг/кг в течение 5 и 7 недель в экспериментальных группах по сравнению с контрольными не обнаружено различий по всем используемым показателям двигательной активности (Табл. 1). Однако через 10 недель введений МФТП наблюдались изменения двух показателей из четырех -пройденный путь и длина шага снизились по сравнению с контролем на 23% и
9
17% соответственно (Табл. 1). Таким образом, через 5 и 7 недель введений МФТП была получена модель досимптомной стадии паркинсонизма, а через 10 недель - модель ранней симптомной стадии.
Биохимические изменения в нигростриатной системе. Через 5 недель введений МФТП содержание ДА в ЧС увеличилось на 40%, а в дальнейшем наблюдалось снижение его содержания - через 7 недель введений МФТП - до уровня контроля, а через 10 недель - на 40% относительно контроля (Рис. 1А).
Табл. 1. Показатели двигательной активности животных через 5, 7 и 10 недель введений МФТП в эксперименте и 0.9% NaCl в контроле.
^Показатель Схемах введенияаа N. МФТП \ |
Пройденный путь (см) |
Число стоек |
Время без движения (сек) |
Длина шага (см) |
||||
0,9%NaCl |
МФТП |
0,9%NaCl |
МФТП |
0,9%NaCl |
МФТП |
0,9%NaCl |
МФТП |
|
5x12 мг/кг |
727,391,7 |
726,6139,0 |
8,01,4 |
11,03,0 |
60,77,2 |
65,69,4 |
4,50,3 |
4,60,3 |
7x12 мг/кг |
1000,2111,7 |
1008,6125,6 |
14,52Д |
10,62,3 |
42,74,6 |
45,47,9 |
4,70,4 |
4,60,5 |
10x12 мг/кг |
989,357,1 |
756,751,6* |
18,82,9 |
14Д1,7 |
51,85,3 |
64,25,9 |
50,2 |
4,30,2* |
*Р<0,05 по сравнению с контролем.
При этом в стриатуме снижение содержания ДА наблюдалось уже через 5 недель введений МФТП и далее продолжало постепенно снижаться (Рис. 1Б). При этом после 10 недель введений достигло порога (80%-го снижения), что, вероятнее всего, и привело к появлению двигательных нарушений. Помимо ДА, в ЧС и в стриатуме на всех стадиях были проанализированы метаболиты ДА -ДОФУК и ГВК, а также моноамины - НА, 5-ГТ и 5-ГИУК. Изменений в содержании НА или 5-ГТ в ЧС ни на одном из сроков обнаружено не было. В стриатуме ни на одной из полученных моделей не было обнаружено достоверных изменений в уровне 5-ГТ, однако на модели ранней досимптомной стадии (через 5 недель введений МФТП) наблюдалось снижение уровня НА на 44%, при этом в дальнейшем его уровень в стриатуме оставался неизменным (Табл. 2). Можно предположить, что уменьшение концентрации НА в стриатуме является показателем нарушения НА-ергической иннервации стриатума на ранней стадии развития патологического процесса в нигростриатной системе. Дальнейшее отсутствие изменений в содержании НА в стриатуме относительно контроля может являться компенсаторным проявлением, так называемой, недофаминергической нейротрансмиссии (Rousselet et al., 2003), наблюдаемой при истощении ДА-ергической нигростриатной системы.
10
Важным функциональным показателем при анализе состояния нигростриатной ДА-ергической системы является соотношение метаболит/медиатор, который в литературе рассматривается как показатель оборота ДА, включающего в себя синтез, распад, выделение и обратный захват
ДА.
Увеличение этого соотношения может интерпретироваться как компенсаторное усиление дофаминовой ейротрансмиссии. В данной работе увеличение соотношения ДОФУК/ДА и ГВК/ДА в стриатуме было обнаружено через 7 недель введений МФТП, т.е. на поздней досимптомной стадии, а через 10 недель введений МФТП - с появлением первых симптомов, данное соотношения значительно снижалось (Табл. 2).
Морфологические изменения e нигростриатной системе. Несмотря на наличие биохимических изменений в ЧС, в компактной части ЧС отсутствовала дегенерация ДА-ергических нейронов через 5, 7 и 10 недель введений МФТП (Рис. 2, ЗА). Также отсутствовали изменения размера (площадь и диаметр) нейронов. При отсутствии дегенерации нейронов в компактной части ЧС, в стриатуме на всех стадиях наблюдалась дегенерация терминалей ДА-ергических аксонов, содержащих оба фермента синтеза ДА - ТГ и ДАА (Рис. ЗБ, 4), при этом размер выживших после введения МФТП терминалей аксонов не изменялся.
Табл. 2. Содержание биогенных аминов и их метаболитов в стриатуме через 5, 7 и 10 недель введений МФТП в опыте и 0.9% NaCl в контроле. Контроль принят за 100%.
11
^\Схема введения ^\^1ФТП Моноаминьг\^^ и метаболиты ^\. |
5x12 мг/кг |
7x12 мг/кг |
10x12 мг/кг |
|||
0,9% NaCl |
МФТП |
0,9% NaCl |
МФТП |
0,9% NaCl |
МФТП |
|
ДОФУК |
1009,3 |
58,33,6* |
1005,9 |
41,62,7* |
1004,3 |
15,22,0* |
ГВК |
10010,8 |
57,13,4* |
1006,4 |
55,44,1* |
1008,2 |
33,91,5* |
ДОФУК/ДА |
1002,5 |
116,24,2 |
1004,8 |
137,76,5* |
1005,2 |
65,28,0* |
ГВК/ДА |
1008,1 |
115,49,6 |
1004,5 |
184,59,7* |
1007,9 |
143,35,1* |
НА |
10013,1 |
56,810,2* |
10018,3 |
11420,8 |
10016,3 |
110,212,5 |
5-ГТ |
1004,5 |
944,2 |
1002,4 |
107,86,6 |
10011,8 |
107,53,5 |
5-ГИУК |
1008,3 |
104,58 |
1002,6 |
107,96,9 |
10011,7 |
93,73,5 |
5-ГИУК/5-ГТ |
1006,5 |
103,62,7 |
1004,5 |
89,13,4 |
10012,1 |
82,44,5 |
*Р<0,05 по сравнению с контролем.
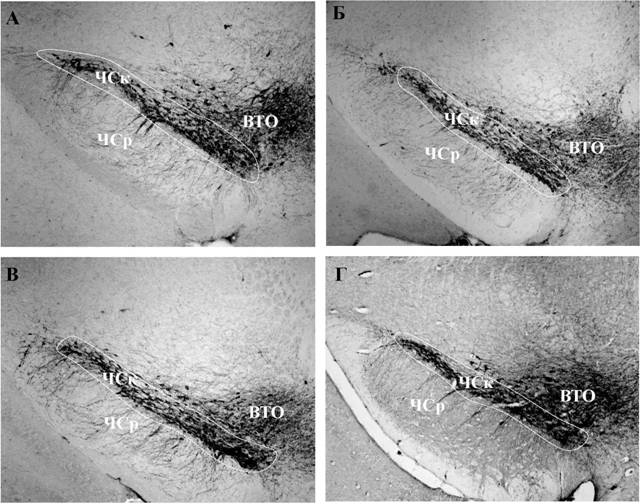
Рис. 2. Световая микроскопия. ТГ-иммунореактивные нейроны в компактной части черной субстанции (ЧСк) в контроле (А) и через 5 (Б), 7 (В) и 10 (Г) недель введений МФТП. Объектив х10. ВТО - вентральная тегментная область; ЧСр ретикулярная часть черной субстанции.
12
Интересно отметить, что помимо терминален аксонов, содержащих оба фермента синтеза ДА, в стриатуме были обнаружены терминали, содержащие только один из ферментов синтеза ДА - либо ТГ, либо ДАА, при этом число моноферментных ДАА-иммунореактивных терминален аксонов уменьшалось после введения МФТП, а число моноферментных ТГ-иммунореактивных -увеличивалось. Аналогичные результаты по изменению иннервации стриатума, а именно увеличение количества моноферментных ТГ-иммунореактивных терминален аксонов, были продемонстрированы и на острых моделях БП. В связи с этим острая модель была выбрана нами как наиболее удобная и быстро-воспроизводимая модель для дальнейшего выяснения происхождения моно ферментных ТГ-иммунореактивных терминален и аксонов стриатума.
Сочетание методов ретроградного мечения нейронов флуоресцентным маркером (трейсером) и иммуногистохимии на ТГ позволило проанализировать все группы нейронов, которые иннервируют стриатум в норме и после введения МФТП. У животных после введения МФТП не было обнаружено групп нейронов, содержащих флуоресцентный трейсер и экспрессирущих ТГ, отличных от контрольных животных. Однако были обнаружены единичные нейроны такого фенотипа в цингулярной коре (Рис. 5А, Б) и гранулярном слое обонятельных луковиц (Рис. 5В, Г).
Таким образом, был сделан вывод, что после введения МФТП может усиливаться регенерация (обратное отрастание) поврежденных ДА-ергических волокон из компактной части ЧС, при этом компенсаторное усиление экспрессии ТГ выражено ярче, чем ДАА, т.к. именно ТГ определяет скорость синтеза ДА. Также не исключается, что может происходить снижение экспрессии ДАА в выживших после действия нейротоксина ДА-ергических терминалях и аксонах стриатума.
Суммируя данные о биохимических и морфологических изменениях в стриатуме можно сделать вывод, что по мере введения МФТП происходит замедление скорости снижения ДА и дегенерации терминалей ДА-ергических аксонов: после первых пяти недель введений МФТП уровень ДА и число терминалей ДА-ергических аксонов в стриатуме снизились на 50% и 47% относительно контроля, тогда как после десяти недель введений - на 80% и 70% относительно контроля (Рис. 6). Таким образом, при хроническом моделировании БП наблюдается медленное и постепенное нарушение функционирования ДА-ергической нигростриатной системы.
Компенсаторные и адаптивные изменения e нигростриатной системе. Замедление скорости снижения ДА в стриатуме может быть связано с увеличением активности ТГ - ключевого фермента синтеза ДА - в выживших терминалях ДА-ергических аксонов стриатума. Действительно, оказалось, что активность ТГ (оцениваемая по накоплению L-ДОФА через 30 минут после введения ингибитора ДАА) в одинаковой степени снижается относительно контроля во всех анализируемых моделях (Рис. 7). С учетом количества дегенерировавших терминалей ДА-ергических аксонов стриатума, можно сделать вывод о компенсаторном увеличении активности ТГ в выживших терминалях ДА-ергических аксонов стриатума, что, вероятнее всего, и привело к отсутствию дальнейшего снижения в них содержания ДА.
Рис. 4. Конфокальнаяаа микроскопия.аа Изображенияаа аксоноваа иаа терминалейаа аксонов стриатума, меченых по ТГ (А, Б, В, Г), ДАА (?', Б', ?', Г') и совмещенные изображения (А", Б", В", Г") в контроле (А, А', А") и после 5 недель (Б, Б', Б"), 7 недель (В, В', В") и 10 недель (Г, Г', Г") введений МФТП. Бар - 10 мкм. Стрелками отмечены терминали разного фенотипа.
Рис. 5. Флуоресцентная микроскопия. Изображения нейронов, меченых трейсером (А, В) и ТГ-иммунореактивных нейронов (Б, Г) в цингулярной коре (А, Б) и гранулярном слое обонятельных луковиц (В, Г) после четырехкратного введения МФТП в дозе 12 мг/кг. Объектив х40. Бар 100 мкм.
14
|
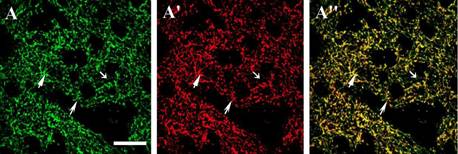
Рис. 4.
|
ЗИ ж;/vw*.' Fir '- |
|
^Х-?^.?;.^-:? |
|
iаа |
B" |
б# ж;..'-а Z?ю Л . . . . -аа ж |
1аа | |
||
Х |
жаа н ''' V'' ж'аа -ж |
|
|
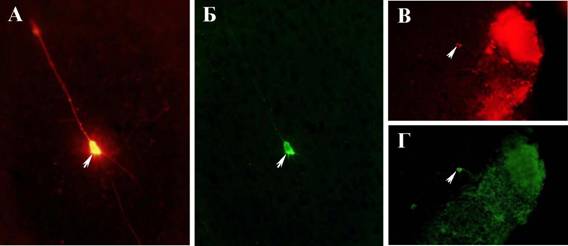
Рис. 5.
В ходе данных экспериментов было показано, что через 10 недель
введений МФТП уровень обратного захвата 3Н-ДА через транспортер ДА в ЧС
и в стриатуме снижается по отношению к контрольным животным на 50% и
57% соответственно (Рис. 11Б). В случае стриатума снижение уровня обратного
захвата 3Н-ДА, вероятно, отчасти отражает дегенерацию терминален ДА-
ергических аксонов. Однако в случае ЧС, где дегенерация тел ДА-ергических
нейронов отсутствовала вовсе, снижение активности транспортера дофамина,
вероятнее всего, является адаптационным механизмом, направленным на
меньшее поступлениеа в нейроны. Нельзя исключить, что на ранних
стадиях развития БП у человека, при наличии эндогенного неиротоксина или экзогенно поступающего неиротоксина, может происходить аналогичная компенсаторная перестройка ДА-ергической нигростриатной системы для сохранения целостности нейрональной популяции.
ВЫВОДЫ
- Саа помощьюаа неиротоксин МФТПаа н мышахаа разработан модель пролонгированного развития БП.
- На модели симптомной стадии БП по сравнению с досимптомной стадией наблюдается:
а) замедление скорости снижения содержания дофамина в стриатуме за счет увеличения активности тирозингидроксилазы в выживших терминалях дофаминергических аксонов;
20
б) увеличение содержания белка тирозингидроксилазы в нейронах
черной субстанции и его уменьшение в аксональных терминалях
стриатума, вероятнее всего, за счет нарушения
аксоплазматического транспорта;
в) отсутствие изменений в экспрессии гена тирозингидроксилазы в
черной субстанции и усиление его экспрессии в стриатуме.
- При моделировании БП увеличивается число тирозингидроксилаза-содержащих волокон в дорзальном стриатуме, однако усиления иннервации стриатума из других областей мозга не обнаруживается.
- На модели ранней симптомной стадии БП происходит уменьшение чувствительности нейронов черной субстанции к нейротоксину МФГГ за счет снижения активности транспортера дофамина, захватывающего МФГГ из межклеточной щели в нейрон.
Статьи в журналах, соответствующих Перечню ВАК и рецензируемых иностранных изданиях:
- Козина Е.А., Хаиндрава В.Г., Кудрин B.C., Кучеряну В.Г., Клодт П.Д., Бочаров Е.В., Раевский КС, Крыжановский Г.Н., Угрюмов М.В. Экспериментальное моделирование функциональной недостаточности нигростриатной дофаминергической системы у мышей // Российский физиологический журнал им. И.М. Сеченова. 2010. Т. 96 (3). С. 270-282.
- Хаиндрава В.Г., Козина Е.А., Кучеряну В.Г., Крыжановский Г.Н., Кудрин B.C., Клодт П.Д., Бочаров Е.В., Раевский КС, Угрюмов М.В. Моделирование преклинической и ранней клинической стадий болезни Паркинсона // Журнал неврологии и психиатрии. 2010. Т. 7. С. 40-47.
- Хаиндрава В.Г., Кудрин B.C., Кучеряну В.Г., Клодт П.Д., Бочаров Е.В., Нанаев А.К., Козина Е.А., Крыжановский Г.Н., Раевский КС, Угрюмов М.В. Экспериментальное моделирование клинической и преклинической стадий болезни Паркинсона // Бюллетень экспериментальной биологии и медицины. 2010. Т. 150 (11). С. 494-497.
- Gurevich I.B., Kozina Е.А., Myagkov A.A., Ugryumov M.V., and Yashina V.V. Automating extraction and analysis of dopaminergic axon terminals in images of frontal slices of the striatum // Pattern Recognition and Image Analysis. Advances in Mathematical Theory and Applications. 2010. V. 20 (3). P. 349-359.
- Ugrumov M.V., Khaindrava V.G., Kozina E.A., Kucheryanu V.G., Bocharov E.V., Kryzhanovsky G.N., Kudrin V.S., Narkevich V.B., Klodt P.M., Rayevsky
21
K.S. and Pronina T.S. Modeling of preclinical and clinical stages of Parkinson's disease in mice//Neuroscience. 2011. V. 181. P. 175-188. 6. Угрюмов M.B., Козина E.A., Хаиндрава В.Г., Кудрин B.C., Кучеряну В.Г., Клодт П.Д., Наркевич В.Б., Бочаров Е.В., Крыжановский Г.Н., Раевский К.С. Моделирование паркинсонизма у мышей при помощи МФТП: от ранней досимптомной до поздней симптомной стадии // Технологии живых систем. 2011. №8. С. 3-13.
Глава в коллективной монографии Угрюмов М.В., Козина Е.А. Модель досимптомной стадии паркинсонизма. В кн.: Нейродегенеративные заболевания: фундаментальные и прикладные аспекты. 2010. С. 223-229.
Труды конференций
- Gurevich I., Koryabkina I., Kozina E., Myagkov A., Niemann H., Ugrumov M., and Yashina V. Method for automatic extraction of dopaminergic neuron terminals on striatum frontal section images // Image Mining Theory and Applications - IMTA 2009 (in conjunction with VISIGRAPP 2009). 2009. P. 30-39.
- Gurevich I., Koryabkina I., Kozina E., Myagkov A., Niemann H., Ugrumov M., and Yashina V. Automated extraction of data for construction of Parkinson's disease experimental model // Pattern Recognition and Information Processing (PRIP). 2009. P. 308-313.
Список сокращений 5-ГИУК, 5-гидроксииндолуксусная кислота; 5-ГТ, 5-гидрокситриптамин, серотонин; L-ДОФА, Ь-3,4-дигидроксифенилаланин; БП, болезнь Паркинсона; ГВК, гомованилиновая кислота; ДА, дофамин;
ДАА, декарбоксилаза ароматических L-аминокислот; ДОФУК, 3,4-дигидроксифенилуксусная кислота; МАО, моноаминоксидаза;
МФТП, 1-метил-4-фенил-1,2,3,6-тетрагидропиридин; МФП+ , 1-метил-4-фенил-пиридин ион; НА, норадреналин; ТГ, тирозингидроксилаза; ЧС, черная субстанция.
22
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]