
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
МОРФОФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ ЛЕЙКОЦИТОВ ТЮЛЕНЕЙ РАЗНОГО ВОЗРАСТА
Автореферат кандидатской диссертации
На правах рукописи
Минзюк
Татьяна Владимировна
МОРФОФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ
ЛЕЙКОЦИТОВ ТЮЛЕНЕЙ РАЗНОГО ВОЗРАСТА
03.03.01 - физиология
Автореферат диссертации на соискание ученой степени
кандидата биологических наук
Петрозаводск - 2012
Работа выполнена в Мурманском морском биологическом институте
Кольского научного центра Российской академии наук
Научный руководитель -
заведующий лабораторией морских млекопитающих,
кандидат биологических наук Кавцевич Николай Николаевич
Официальные оппоненты:
Мейгал Александр Юрьевич - доктор медицинских наук, профессор Ц
Петрозаводский государственный университет,
профессор кафедры физиологии человека и животных;
Олейник Евгения Константиновна - доктор биологических наук Ц
Институт биологии Карельского научного центра
Российской академии наук, руководитель группы иммунологии.
Ведущая организация - Мурманский государственный
гуманитарный университет
Защита состоится л а30а аа маяаа 2012 г.а в 12 часов на заседании диссертационного совета ДМ 212.087.02 при Карельской государственной педагогической академии по адресу: 185035, Республика Карелия,
г. Петрозаводск, ул. Пушкинская, 17.
С диссертацией можно ознакомиться в библиотеке Карельской государственной педагогической академии.
Автореферат разослана л____ _________________ 2012 г.

Ученый секретарь диссертационного совета А.И. Малкиель
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Изучение закономерностей развития реактивности лейкоцитов является одним из важных направлений исследования механизмов резистентности организма (Бахов, 2000; Долгушин, Бухарин, 2001; Levy, 2004; Соколова, 2005; Антонишкис, 2006; Пинегин, Маянский, 2007; Будыка и др., 2009; Frouin et al., 2010; Cederlund et al., 2011).
У морских млекопитающих имеются особенности строения и функционирования кроветворной системы, обусловленные длительной эволюцией в водной среде, не содержавшей патогенных микроорганизмов (Cavagnolo, 1979; Сыкало, 1989; Соколова, Денисенко, 2006). В то же время, механизмы специфической и неспецифической резистентности водных млекопитающих исследованы недостаточно для оценки их эффективности по сравнению с таковыми наземных животных.
Серый тюлень (Halichoerus grypus) - охраняемый вид ластоногих, включенный в Красную книгу. Гренландский тюлень (Pagophilus groenlandicus) и тюлень-хохлач (Cystophora crystata) являются объектами коммерческого зверобойного промысла. Интенсивное исследование гематологических и биохимических показателей крови хохлача проводилось только в Канаде (Boily et al., 2006). В России такие работы, как и посвященные изучению крови серого тюленя, единичны. Гренландский тюлень, в связи с прекращением его промысла в России, также стал труднодоступным объектом исследования.
Изучение резистентности организма данных животных, особенно в ранние периоды постнатального онтогенеза, необходимо для выявления причин естественной смертности в стаде, оценки перспектив развития популяций этих морских млекопитающих. Гематологические показатели диких и плененных животных могут быть использованы в ветеринарной практике, в оценке хода реабилитации и при длительном содержании животных в условиях неволи.
У новорожденных млекопитающих иммунная система находится на стадии формирования. В период молочного питания временной защитой от патогенных микроорганизмов для щенков тюленей выступают материнские антитела, поступающие через молозиво от самки (King et al., 1994; Ross et al., 1993, 1994). Поэтому иммуннокомпетентность щенков в первые недели постнатального онтогенеза зависит от длительности периода молочного питания (Ross et al., 1994). В период голодания после молочного вскармливания и последующего перехода к самостоятельному питанию противоинфекционную защиту организма детенышей тюленей обеспечивают реакции врожденного неспецифического клеточного иммунитета. В связи с этим важно изучить параметры различных типов лейкоцитов крови для понимания функционального значения их изменений в раннем постнатальном онтогенезе тюленей.
Определение цитохимических параметров лейкоцитов позволяет идентифицировать группы клеток с различными иммунологическими функциями, оценить их функциональную активность. Имеющейся в настоящее время в литературе информации о формировании специфических и неспецифических иммунных реакций у морских млекопитающих, особенно в ранние периоды постнатального онтогенеза, недостаточно для суждения об уровне резистентности этих животных. В связи с этим актуально исследование морфофункциональных свойств лейкоцитов крови у тюленей, как в природных популяциях, так и в условиях неволи на разных этапах постнатального онтогенеза.
Исследование морфофункциональных особенностей лейкоцитов у морскиха млекопитающих даст возможность оценить процессы адаптации и устойчивости в естественной среде обитания, а также является актуальным для оценки состояния здоровья животных при содержании в условиях неволи.
Большой теоретический интерес для сравнительной физиологии представляют параметры клеток крови водных млекопитающих, в особенности связанные с обеспечением резистентности этих животных к воздействию неблагоприятных факторов.
Цель и задачи исследования. Целью настоящей работы являлась оценка морфофункциональных особенностей внутриклеточных компонентов бактерицидных систем лейкоцитов и ядрышкообразующих районов лимфоцитов периферической крови у тюленей разного возраста.
Для достижения цели были поставлены следующие задачи:
- Определить особенности клеточного состава крови тюленей.
- Рассчитать гематологические индексы у тюленей разного возраста.
- Определить активность миелопероксидазы и содержание бактерицидного катионного белка в лейкоцитах тюленей.
- Исследовать морфометрические параметры окрашенных серебром районов организаторов ядрышка в лимфоцитах тюленей разного возраста.
Научная новизна. Впервые проведено морфологическое и цитохимическое исследование клеточного состава периферической крови щенков тюленя-хохлача, завершивших молочное вскармливание. Впервые определены лейкоцитарные индексы серых и гренландских тюленей в различные периоды раннего постнатального онтогенеза. Впервые определено содержание бактерицидных белков лейкоцитов у тюленей в разные возрастные периоды. Впервые проведена оценка функционального состояния лимфоцитов периферической крови ластоногих по морфометрическим параметрам окрашенных серебром районов организаторов ядрышка.
Практическая и теоретическая значимость работы. Результаты гематологических и цитохимических исследований могут быть использованы для оценки состояния здоровья тюленей, как в естественной среде обитания, так и в ходе реабилитации, лечения и длительного содержания в условиях неволи. Полученные в работе новые данные расширяют представления о роли показателей клеточного состава крови и внутриклеточных компонентов лейкоцитов в формировании специфических и неспецифических клеточных адаптационных реакций у морских млекопитающих.
Положения, выносимые на защиту.
- Наибольшие изменения в клеточном составе крови тюленей происходят в первые 3-4 месяца постнатального онтогенеза. Сроки наступления физиологического перекреста лейкоцитарной формулы крови у серого, гренландского тюленей и тюленя-хохлача различны.
- У хохлача, серого и гренландского тюленей очень низкое содержание катионного белка по сравнению с наземными животными.
- В лимфоцитах тюленей до 3-4 месяцев уровень синтеза рРНК выше, чем у взрослых животных.
Апробация работы. Материалы работы доложены и обсуждались на международных научно-практических конференциях по морским биотехническим системам и освоению морских и наземных экосистем в условиях арктического и аридного климата (Ростов-на-Дону, 2008 и 2011), II международной конференции "Актуальные проблемы экологической физиологии, биохимии и генетики животных" (Саранск, 2009); на международном симпозиуме по физиологии и патологии млекопитающих, введенных в зоокультуру (Петрозаводск, 2009), на XXVI, XXVII, XXVIII и XXIX научных конференциях молодых учёных (Мурманск, 2008, 2009, 2010 и 2011), международной конференции студентов, аспирантов и молодых ученых Ломоносов - 2009 (Москва, 2009), V-VI конференциях "Морские млекопитающие Голарктики" (Одесса, 2008; Калининград, 2010).
Публикации. По теме диссертации опубликованы 24 научные работы, в том числе 5 статей ва рецензируемых журналах, рекомендованных ВАК РФ.
Структура и объем работы. Диссертация изложена на 124 страницах машинописного текста, содержит 17 таблиц и 38 рисунков. Работа состоит из введения, обзора литературы, описания материалов и методов исследования, 3-х глав результатов собственных исследований, заключения, выводов и списка литературы, который включает 228 наименований, из них 119 иностранных.
Материалы и методы исследования
Объекты исследования. Гренландский тюлень (Pagophilus groenlandica Erxleben, 1777) следующих возрастных групп: 1. бельки, возраст 1 неделя (n=38), 2а. серки, 1-1.5 месяца, "нормальные" - ахорошо упитанные детеныши весом 30-35 кг (n=27), 2б. серки, 1-1.5 месяца, "заморыши" - весом 10-15 кг (n=36), 3. серки, 3-4 месяца (n=6), 4. взрослые животные (n=10). В условиях неволи получали кровь от 4 животных, содержавшихся в аквакомплексе экспериментального морского полигона ММБИ в период 2001-2004гг.
Серый тюлень (Halichoerus grypus Fabricius, 1791): 1. новорожденные (n=8), 2. питающиеся молоком, 2-3 недели (n=6), 3. завершившие молочное питание, 1-1.5 месяца (n=12), 4. самостоятельно питающиеся рыбой щенки, 3-4 месяца (n=6), 5. взрослые животные, 1 год (n=4) и 3.5 года (n=2).
Тюлень-хохлач (Cystophora cristata Erxleben, 1777), 1-1.5 месяца (n=13).
Методы исследования. Кровь у тюленей брали из внутрипозвоночной вены (Geraci, Smith, 1975), от хохлачей во время промысла - из правого желудочка сердца в шприц с гепарином. Мазки крови изготавливали общепринятым способом (Предтеченский, 1960).
Морфологию лейкоцитов изучали на препаратах, фиксированных в метаноле 5 минут и окрашенных по Романовскому. Для выявления катионного белка фиксированные препараты помещали в 0.1% раствор прочного зеленого FCF (рН 8.1-8.2) на 1 час (Бутенко и др., 1974), клеточные ядра докрашивали нейтральным красным. Миелопероксидазу в лейкоцитах выявляли по методу Д. Кваглино (Лецкий, 1973), помещая фиксированные в смеси формалина и этанола мазки в спиртовой раствор бензидина и перекиси водорода на 30 мин. Ядра докрашивали по Романовскому. Белки районов организаторов ядрышек окрашивали азотнокислым серебром по методике У. Хоуэлла и Д. Блэка (1980). Мазки погружали на 30 мин в свежеприготовленную смесь из 2 частей 50% нитрата серебра и 1 части 1% муравьиной кислоты в 2%-ном растворе желатина.
Морфометрические параметры внутриклеточных компонентов лейкоцитов (количество, диаметр, площадь, округлость) определяли с использованием микроскопа Axio Imager M1, оснащенного цифровой видеокамерой AxioCam и программным обеспечением AxioVision (фирмы Zeiss). Препараты изучали, используя масляную иммерсию (объектив ?100, окуляр ?10). Мерой активности ядрышек служила относительная площадь окрашенных серебром районов организаторов ядрышка (отношение площади ядрышкообразующего района к площади ядра лимфоцита). При определении содержания катионного белка и миелопероксидазы в гранулоцитах крови вычисляли средний цитохимический коэффициент. Для определения адаптационных состояний тюленей рассчитывали лейкоцитарные индексы (Гаркави, 1990; Мустафина и др., 1999; Сперанский и др., 2004; Островский и др., 2007): адаптационный индекс, модифицированный лейкоцитарный индекс интоксикации, ядерный индекс интоксикации, реактивный ответ нейтрофилов, индекс сдвига лейкоцитов, лимфоцитарно-гранулоцитарный индекс, индекс соотношения нейтрофилов и моноцитов.
Статистическую обработку результатов проводили методами вариационной статистики с использованием программ Microsoft Excel Windows XP и Statistica 6.0. Рассчитывали среднюю и ошибку средней (Mm), коэффициент вариации (С.V.), показатели асимметрии (As) и эксцесса (Ex). Уровень сходства гистограмм распределения клеток по морфометрическим параметрам определяли при помощи критерия ?2. Для оценки достоверности различий между средними величинами использовали критерий Стьюдента, различия при p<0.05 рассматривались как статистически значимые.
Результаты исследования
Клеточный состав крови тюленей разного возраста
Показатели лейкоцитарной формулы существенно отличались в крови у тюленей разных возрастных групп (рис. 1). У новорожденных серых тюленей и в первые 1-1.5 месяца у гренландских в крови встречаются юные клетки: метамиелоциты и нормоциты. Во всех группах тюленей встречаются палочкоядерные нейтрофилы (1-4%).
У хохлача и серого тюленя в 1-1.5 месяца, а у гренландского в 2-3-х недельном возрасте (рис. 1) происходит уравнивание количества сегментоядерных нейтрофилов и лимфоцитов. Физиологический перекрест на ранних этапах постнатального онтогенеза у тюленей, наиболее вероятно, обусловлен интенсивной пролиферацией лимфоидных клеток, связанной с развитием системы специфического иммунитета животных. А высокий уровень сегментоядерных нейтрофилов в первые дни и месяцы жизни является приспособлением, обеспечивающим неспецифическую защиту организма от инфекций (Алексеев, 1998).
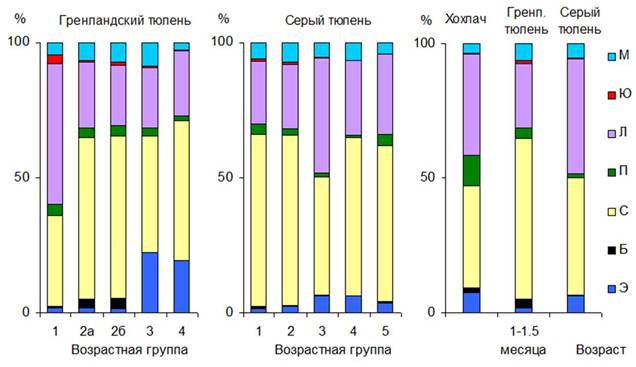
Рис. 1. Лейкоцитарная формула крови тюленей: М - моноциты, Ю - юные клетки, Л - лимфоциты, П - палочкоядерные нейтрофилы, С - сегментоядерные нейтрофилы, Б - базофилы, Э - эозинофилы.
В крови хохлачей 1-1.5 месяцев в большом количестве присутствуют эозинофилы и палочкоядерные формы лейкоцитов. Эозинофилия в периферической крови млекопитающих наблюдается при аллергических реакциях разного происхождения. Роль аллергенов могут играть вещества, поступающие с воздухом и пищей (особенно в период начала самостоятельного питания). Как одну из вероятных причин эозинофилии крови китообразных рассматривают гельминтозы (Богданова, Лебедев, 1971; Ridgway, 1972; Greig et al., 2010).
Отдельные показатели лейкоцитарной формулы не дают целостного представления о реакциях организма на воздействие различных факторов. Поэтому в исследованиях человека и животных применяют лейкоцитарные индексы, позволяющие достоверно диагностировать и прогнозировать изменения состояния организма. "Нормальные" значения лейкоцитарных индексов взрослых людей мы использовали в качестве ориентировочных при определении адаптационного статуса тюленей, поскольку показатели лейкоцитарной формулы крови взрослых тюленей близки к таковым у человека.
Резкие изменения индексов у щенков серых тюленей, завершивших молочное питание, и у бельков гренландских тюленей (рис. 2, 3) обусловлены характерным для этих периодов уравниванием количества лимфоцитов и нейтрофилов. Тем не менее, значимыми и информативными при определении адаптационного статуса тюленей могут быть: модифицированный лейкоцитарный индекс интоксикации, реактивный ответ нейтрофилов (рис. 3) и индекс соотношения нейтрофилов и моноцитов. Уровни этих показателей у новорожденных тюленей и животных других возрастных групп достоверно различаются (p<0.01) и существенно изменяются при переходе в следующий период развития. Вероятно, эти различия не связаны с инфекционными и интоксикационными процессами в организме животных, а обусловлены возрастными особенностями клеточного состава крови.
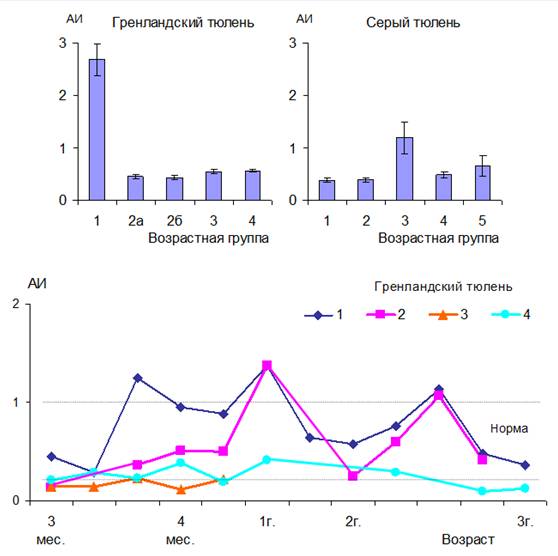
Рис. 2. Адаптационный индекс (АИ) у тюленей разного возраста.
Использование модифицированного лейкоцитарного индекса интоксикации, реактивного ответа нейтрофилов (рис. 3), адаптационного индекса (рис. 2) и индекса соотношения нейтрофилов и моноцитов эффективно при оценке адаптационных возможностей ослабленных животных в естественной среде обитания, в ходе их реабилитации и адаптации к условиям неволи. Согласно системе оценок адаптационных возможностей организма Л.Х. Гаркави (1990), у всех гренландских тюленей в первые дни пребывания в неволе наблюдается реакция стресса. Это подтверждается низким относительным количеством лимфоцитов (менее 20%). У животного №3, которое было истощено и в дальнейшем погибло, адаптационный индекс не повышался до нормальных значений.
Вероятно, для взрослых тюленей реактивный ответ нейтрофилов (рис. 3) в норме значительно ниже, чем для человека. Истощенные же щенки гренландских тюленей, согласно полученным результатам, подвергались интоксикации средней тяжести как в естественной среде обитания, так и в условиях неволи.
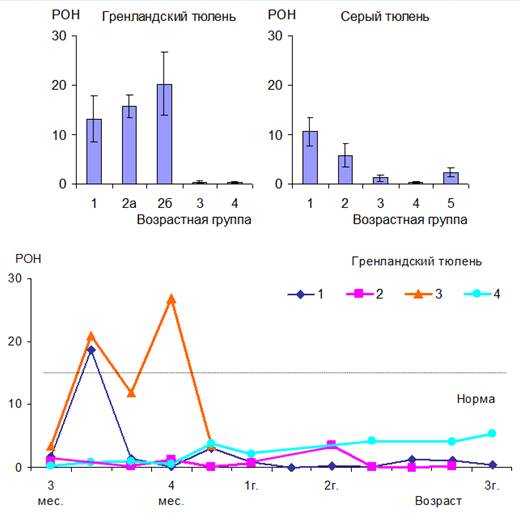
Рис. 3. Реактивный ответ нейтрофилов (РОН) у тюленей разного возраста.
В гематологических исследованиях ластоногих необходимо применение простых и в то же время чувствительных и информативных показателей для оценки их физиологического и функционального состояния, поскольку контакт с этими водными животными на суше должен быть максимально редким и щадящим. Поэтому в следующем разделе работы нами исследованы цитохимические параметры лейкоцитов тюленей.
Цитохимическая характеристика лейкоцитов крови тюленей
Цитохимические исследования по определению содержания миелопероксидазы и катионного белка в гранулоцитах, а также подсчет числа БГЛ в крови имеют значение при оценке уровня неспецифической резистентности организма животных (Рулева и др., 2007). Главными факторами бактерицидной активности лейкоцитов являются кислородзависимая МПО и кислороднезависимые неферментные КБ. Недостаточность КБ и МПО в гранулоцитах приводит к резкому снижению неспецифической резистентности организма (Пигаревский, 1978).
В гранулоцитах всех исследованных морских млекопитающих наблюдается гранулярное распределение катионного белка (рис. 4). Средняя площадь одной гранулы у тюленей составляет 0.26 мкм2 (у человека 0.30 мкм2). С возрастом происходит увеличение среднего числа окрашиваемых прочным зеленым гранул на клетку: у серого тюленя - от 95 (новорожденные) до 140 (взрослые), у гренландского тюленя - от 75 (бельки) до 155 (взрослые).

Рис. 4. Катионный белок в гранулоцитах гренландского (А), серого (В) и тюленя-хохлача (Б).
У щенков серого тюленя до 1.5 месяцев и гренландского - до 3-х число лейкоцитов, в которых выявляются катионные белки, очень низкое (2-4.5%) (рис. 5). При этом у 19% серок гренландского тюленя (возраст 1-1.5 мес.) гранулоциты, содержащие катионные белки, не выявлены.

Рис. 5. Содержание катионного белка в гранулоцитах тюленей.
Достоверное увеличение КБ происходит к 3-4 месяцам (р<0.01), это проявляется в увеличении числа КБ+лейкоцитов и содержащихся в них гранул. Это, по-видимому, обусловлено сложными физиологическими перестройками, связанными с голоданием и последующим переходом щенков на самостоятельное питание рыбой, и свидетельствует об увеличении противоинфекционной неспецифической резистентности организма тюленей. Содержание КБ у взрослых гренландских тюленей в 2 раза выше, чем у серых (рис. 5).
У тюленей на всех исследованных возрастных этапах очень низкое содержание КБ по сравнению с другими видами животных и человеком (рис. 6).
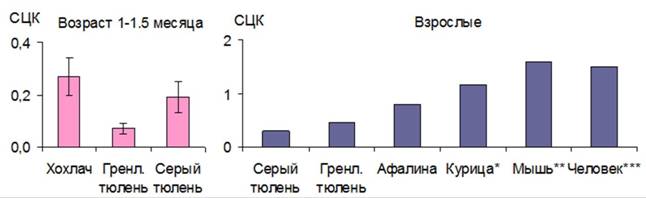
Рис. 6. Содержание катионного белка в гранулоцитах тюленей разных видов животных (курица, мышь и человек по: *Клетикова, 2010, **Будыка и др., 2009, ***Стойко, Ермаков, 2004).
Изучение гренландских тюленей, содержащихся в условиях неволи в течение длительного времени, показало индивидуальные различия между животными по количеству КБ+лейкоцитов (рис. 7). В возрасте 3-4-х месяцев у тюленей №2 и №4 содержание КБ+лейкоцитов повышалось до 0.5 у.е., что близко к показателям животных в естественной среде обитания. В данный возрастной период тюлени переходят на самостоятельное питание рыбой, как в природных условиях, так и в неволе. Самое низкое содержание катионного белка наблюдалось у животного №3, которое было истощено и впоследствии погибло. У взрослых плененных гренландских тюленей содержание бактерицидного катионного белка в 2.6 раза ниже (СЦК=0.170.03), чем у диких животных (СЦК=0.450.06).

Рис. 7. Возрастные изменения содержания катионного белка в гранулоцитах гренландских тюленей в условиях неволи.
Таким образом, у тюленей в начальные периоды постнатального онтогенеза очень низкое содержание катионного белка в гранулоцитах. С 3-х месячного возраста у гренландского тюленя и с 1-1.5 месяцев у хохлача и серого тюленей происходит интенсивное увеличение числа КБ+лейкоцитов и содержащихся в них лизосомных гранул. При этом показано, что резкое возрастание показателей фагоцитоза у щенков серых и гренландских тюленей происходит несколько раньше, в период молочного питания (Frouin et al., 2010).
Известно, что максимальный бактерицидный эффект достигается при совместном действии катионного белка и миелопероксидазы (Пигаревский, 1978). При этом показано, что микробицидное действие КБ гранулоцитов человека не зависит от МПО (Odeberg, Olsson, 1975). МПО участвует в галогенизации белков бактерии (Рулева и др., 2007; Рязанцева, 2009), а также, являясь ферментом АОС, обеспечивает защиту организма от окислительного стресса (Галанцев и др., 1999; Молчанов и др., 1999; Коваленко и др., 2004). Предполагают, что ферменты АОС, в частности пероксидазы, участвуют в освобождении эндогенного молекулярного кислорода для поддержания аэробных процессов при апноэ (Молчанов и др., 1999; Ноздрачев и др., 2000; Баранова и др., 2010).
МПО при окрашивании гранулоцитов у серых тюленей характеризуется диффузно-гранулярным распределением (рис. 8). Средний размер зерен окрашенного продукта цитохимической реакции на МПО в нейтрофильных гранулоцитах составляет 0.42 мкм2, в эозинофильных гранулоцитах они крупнее - 0.55 мкм2.

Рис. 8. Миелопероксидаза в гранулоцитах серых тюленей.
У ныряющих млекопитающих различных видов (норки, ондатры, дельфины) активность ферментов АОС выше, чем у наземных животных (Молчанов и др., 1999). Нами показано достоверное снижение показателей СЦК с возрастом (рис. 9): от 2.51 (новорожденные) до 1.91 (взрослые). У взрослого серого тюленя, имеющего опыт погружений под воду, активность фермента самая низкая.
Вероятно, на ранних этапах онтогенеза тюленей ведущую роль в регуляции активности миелопероксидазы играет микрофлора среды, в которую попадают детеныши после рождения. У щенков тюленей, длительное время (1-1.5 месяцев) находящихся на суше, одной из основных функций МПО может быть обеспечение неспецифических защитных реакций лейкоцитов. Высокая активность МПО у новорожденных обусловлена, вероятно, жизненно важной ролью этого фермента в АОС в период внутриутробного развития. После рождения возрастает значение бактерицидного действия этого фермента, которое дополняется повышающейся с возрастом активностью неферментного компонента бактерицидной защиты, лизосомных катионных белков.

Рис. 9. Содержание катионного белка и миелопероксидазы в гранулоцитах серых тюленей (мышь и человек по: *Будыка и др., 2009; **Палагнюк, 2003 (МПО); Стойко, Ермаков, 2004 (КБ)).
Одним из важнейших компонентов врожденного клеточного иммунитета являются естественные киллерные клетки (ЕКК), разрушающие опухолевые и инфицированные вирусами клетки. ЕКК рассматривают как первую линию обороны системы иммунитета, менее специфичную, чем индуцированный иммунитет, но быстрее реагирующую. Все ЕКК по своей морфологии "большие гранулосодержащие лимфоциты". БГЛ тюленей представляют собой клетки крупного или среднего размера, в цитоплазме содержатся мелкие азурофильные гранулы (0.1-0.2 мкм) вишневого цвета (рис. 10).

Рис. 10. Лимфоциты тюленей: а - малый лимфоцит; б-в - БГЛ с узким ободком цитоплазмы; г-е - "типичные" БГЛ тюленей.
Реакции специфического иммунитета у новорожденных животных еще не сформированы в достаточной степени, поэтому роль БГЛ в первые дни и недели жизни может быть значительной и их отсутствие (или низкое число) в крови расценивается "как нежелательное" (Потапов и др., 1992). Содержание БГЛ в крови гренландских тюленей на всех возрастных этапах выше, чем у хохлача (в 1-1.5 месяца 2.10.6%) и серого тюленей (рис. 11). Максимальное повышение количества БГЛ наблюдается в период завершения молочного питания и у серых и гренландских тюленей. Причем, это повышение происходит не только у нормальных, но и у истощенных ("заморышей") щенков гренландского тюленя, чье физиологическое состояние, очевидно, значительно отличается от состояния первых.
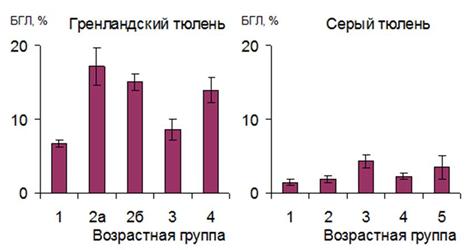
Рис. 11. Относительное количество БГЛ у тюленей разного возраста.
Большие гранулярные лимфоциты широко представлены в филогенетическом ряду. Естественная клеточная цитотоксичность обнаружена у рыб, амфибий, рептилий и птиц, а также земляного червя, морского ежа, асцидии, губок (Галактионов, 2005). В крови здоровых людей содержится 2-12% БГЛ, у мышей - до 9.1%, у крыс - до 9.1% (Зак, Бутенко, 1985). Таким образом, содержание БГЛ в крови гренландских тюленей выше, чем у многих наземных млекопитающих. Это может свидетельствовать о большей роли клеточно-опосредованной цитотоксичности у этих животных.
Определение содержания миелопероксидазы и катионного белка в гранулоцитах, а также подсчет числа БГЛ в крови дает информацию о формировании неспецифических защитных реакций у тюленей разного возраста. Определение функциональной активности организаторов ядрышка в лимфоцитах позволяет судить о потенциальных возможностях клеток синтезировать белок. В следующем разделе мы исследовали морфометрические параметры районов организаторов ядрышка, отражающие уровень синтеза рРНК в лимфоцитах тюленей.
Морфометрические параметры районов организаторов ядрышка в лимфоцитах тюленей
Окрашивание белков районов организаторов ядрышка, участвующих в процессах биосинтеза и созревания пре-рРНК (Челидзе, Черкезия, 2005; Жарская, Зацепина, 2007; Biggiogera et al., 2001;Lam et al., 2005), позволяет выявлять изменения функциональной активности ядрышка клеток. Увеличение числа и размеров, изменение формы окрашенных серебром ЯОР (AgЯОР), наблюдается в клетках с высокой пролиферативной активностью (Sirri et al., 2000; Derenzini, Trerе, 2001).
AgЯОР в лимфоцитах тюленей имеют преимущественно округлую или слегка вытянутую с небольшими неровностями контуров форму. В период с 1-1.5 до 3-4 месяцев у щенков серого и гренландского тюленей увеличивается число неправильных по форме AgЯОР: вытянутых, с заостренными углами (рис. 12).
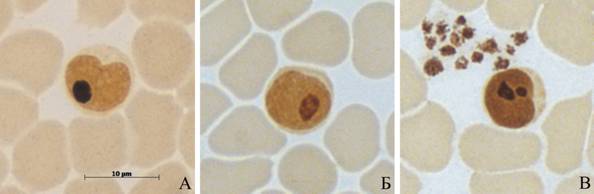
Рис. 12. AgЯОР в лимфоцитах тюленей.
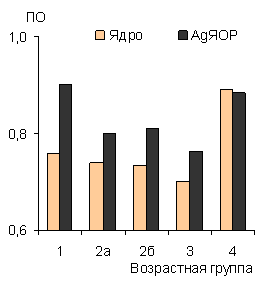
Рис. 13. Показатель округлости (ПО) ядра и AgЯОР в лимфоцитах гренландского тюленя. |
Показатель округлости AgЯОР снижается у щенков с 1-1.5 месяцев, и возрастает у взрослых животных (рис. 13). Известно, что показатель формы идеально круглых объектов равен 1. Все объекты со значениями меньше 1 обладают неправильной формой. Вероятно, в возрасте от 1 до 4 месяцев у щенков тюленей высокий полиморфизм клеточных структур лимфоцитов по сравнению с взрослыми животными свидетельствует о высокой пролиферативной активности лимфоцитов. |
Изменения в морфологии ядра интерфазных лимфоцитов отмечены при лейкозах у коров, также у них наблюдается увеличение "атипичных" по форме AgЯОР (Косякова, 2010). В опухолевых клетках проявляется выраженный ядрышковый полиморфизм (Мамаев, 1991), при этом AgЯОР раковых клеток имеют более неправильные формы, чем контрольные клетки (Simeonov, Simeonova, 2006).
Максимальное число ядрышкообразующих районов в интерфазных клетках определяется числом ядрышкообразующих хромосом, у исследуемых нами морских млекопитающих составляет 4 (Arnason, 1974; Arnason et al., 1993, 1995). У серого тюленя лимфоциты с 4-мя AgЯОР встречаются в 0-4% случаев, у гренландского же тюленя максимальное их число было выявлено только у животных в условиях неволи (не более 1-2%). Не все организаторы ядрышка визуализируются в лимфоцитах, т.к. некоторые из них либо вступают в ассоциации (Goessens, 1984; Сабанеева, 1989; Мухарьямова, Зацепина, 1993), либо не являются активными (Wachtler et al., 1986; Shadley, 1994). У всех тюленей преобладают лимфоциты с одним AgЯОР (73.5-90.0%), и только у хохлача в возрасте 1-1.5 месяцев - с двумя AgЯОР (46.962.59%).
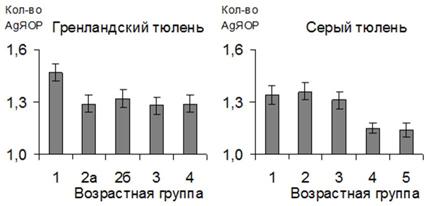
Рис. 14. Количество AgЯОР в лимфоцитах тюленей.
У гренландских тюленей в 1.5 месяца, у серых - в 3-4 месяца наблюдается увеличение количества лимфоцитов с одним AgЯОР (рис. 14). При этом щенки серых тюленей, завершившие молочное вскармливание, но еще не питающиеся рыбой, а также бельки гренландских тюленей характеризуются повышенным относительным количеством лимфоцитов по сравнению с другими возрастными группами (р<0.05).
Относительный размер AgЯОР (SAgЯОР:SЯ) от рождения и до начала самостоятельного питания у щенков тюленей выше, чем у взрослых животных (табл. 1, рис. 15).
Таблица 1
Статистические параметры распределений лимфоцитов тюленей по величине отношения площади AgЯОР к площади ядра
Группы тюленей |
Мm |
As |
Ex |
C.V. |
Серый тюлень |
||||
1 |
0.1270.001 |
0.707 |
0.887 |
24.7 |
2 |
0.1340.001 |
0.275 |
0.001 |
28.1 |
3 |
0.1220.001 |
0.351 |
0.404 |
25.2 |
4 |
0.1040.001 |
0.723 |
1.043 |
25.9 |
5 |
0.0870.006 |
1.484 |
3.975 |
34.3 |
Гренландский тюлень |
||||
1 |
0.0870.001 |
0.772 |
0.634 |
36.1 |
2а |
0.0950.001 |
0.599 |
0.525 |
31.0 |
2б |
0.0920.001 |
0.722 |
0.735 |
33.2 |
3 |
0.1040.001 |
0.503 |
0.202 |
26.6 |
4 |
0.0820.001 |
0.726 |
1.786 |
35.9 |
Примечание. Мm - средняя и ошибка средней; As - асимметрия; Ex - эксцесс; C.V. - коэффициент вариации
Взрослые животные характеризуются высоким показателем эксцесса (табл. 1), т.е. в периферической крови у них преобладают лимфоциты, сходные по функциональному состоянию. В отличие от взрослых животных щенкам тюленей свойственно наличие больших групп лимфоидных клеток, существенно различающихся по интенсивности синтетических процессов (сравнение по ?2, p<0.05), т.е., согласно полученным результатам, в первые 3-4 месяца жизни тюленей воздействие антигенов внешней среды (а также внутренних факторов, в частности, гормонов) на кроветворную систему тюленей наиболее значительно. Относительная площадь организаторов ядрышка у щенков серых тюленей в постнатальном периоде развития выше, чем у гренландских тюленей (табл. 1, рис. 15).

Рис. 15. Распределение лимфоцитов по величине SAgЯОР:SЯ у тюленей разных возрастных групп.
Наибольшее число AgЯОР в одном лимфоците у тюленей наблюдается в первые месяцы адаптации к неволе и достоверно снижается на первом году жизни, при этом относительный размер AgЯОР значительно не меняется (рис. 16).

Рис. 16. Возрастные изменения числа AgЯОР и величины SAgЯОР:SЯ у гренландских тюленей.
Таким образом, синтез рРНК в клетках формирующейся лимфоидной системы детенышей тюленей более интенсивен, чем у взрослых животных, что проявляется в большем числе и относительном размере AgЯОР, а также в увеличении числа неправильных по форме организаторов ядрышка и преобладании субпопуляций лимфоцитов, различных по своему функциональному состоянию.
Наиболее высокие показатели числа AgЯОР на клетку наблюдаются у серых тюленей от рождения до 1-1.5 месяцев (щенки завершили молочное питание), у гренландских тюленей - у бельков. Эти данные согласуются с показателями лейкоцитарной формулы: у серых тюленей в 1-1.5 месяца и у гренландских тюленей в первый месяц жизни происходит возрастание числа лимфоцитов и уравнивание последних с числом сегментоядерных нейтрофилов. Следовательно, высокое количество активных AgЯОР в лимфоцитах крови подтверждает, что в период физиологического перекреста происходит усиление транскрипции рибосомных генов и пролиферативных процессов в организме животных.
Таким образом, первые 3-4 месяца постнатального онтогенеза тюленей характеризуются наиболее значительными изменениями в клеточном составе крови, активизацией бактерицидных систем организма и повышением функциональной активности организаторов ядрышка лимфоцитов. Это связано с активно идущими процессами становления костномозгового кроветворения и развития клеточных защитных механизмов.
ВЫВОДЫ
- В периферической крови щенков хохлача, серого и гренландского тюленей встречаются низкодифференцированные формы лейкоцитов и эритроцитов. Физиологический перекрест лейкоцитарной формулы крови у гренландских тюленей происходит в возрасте 2-3 недель, у хохлача и серого тюленей - в 1-1.5 месяца. У гренландских тюленей в возрасте 3-4 месяцев и у взрослых животных наблюдается высокое содержание эозинофилов (19-23%). Количество больших гранулярных лимфоцитов у гренландских тюленей значительно выше, чем у хохлача и серого тюленей.
- Наибольшие изменения лейкоцитарных индексов происходят у тюленей в периоды физиологического перекреста лейкоцитарной формулы крови, что не связано с инфекционными и интоксикационными процессами в организме. В первые месяцы адаптации к условиям неволи существенно меняются значения адаптационного индекса, модифицированного лейкоцитарного индекса интоксикации, реактивного ответа нейтрофилов и индекса соотношения нейтрофилов и моноцитов.
- Содержание катионного белка у серых тюленей повышается в возрасте 1-1.5 месяцев, у гренландских тюленей - в 3-4 месяца (увеличивается как количество КБ+лейкоцитов, так и число КБ+гранул). У взрослых тюленей в условиях неволи содержание КБ в 2.6 раза ниже, чем у диких животных.
- В процессе раннего постнатального онтогенеза серых тюленей активность миелопероксидазы в гранулоцитах снижается. При этом бактерицидная роль миелопероксидазы у тюленей значительно выше, чем катионного белка.
- Активность организаторов ядрышка в лимфоцитах детенышей тюленей выше, чем у взрослых животных, что проявляется в большем количестве и относительном размере AgЯОР, высоком полиморфизме организаторов ядрышка. В первые месяцы адаптации к условиям неволи у всех гренландских тюленей наблюдается высокое количество AgЯОР. Истощенные животные в естественной среде обитания и в условиях неволи отличаются низкими показателями относительного размера AgЯОР.
ОСНОВНЫЕ ПУБЛИКАЦИИ ПО ТЕМЕ ДИССЕРТАЦИИ
Статьи, опубликованные в рецензируемых научных журналах
- Кавцевич Н.Н., Минзюк Т.В. Клеточный состав крови серых тюленей различного возраста // ДАН. - 2010. - Т. 432, № 4. - С. 552Ц555.
- Минзюк Т.В., Кавцевич Н.Н. Новые данные о клеточном составе крови тюленя хохлача // ДАН. - 2010. - Т. 435, № 5. - С. 714Ц717.
- Кавцевич Н.Н., Минзюк Т.В. Лейкоцитарные индексы и активность организаторов ядрышка лимфоцитов щенков серых тюленей // Вестник ЮНЦ. - 2010. - Т. 6, № 4. - С. 76Ц83.
- Кавцевич Н.Н., Минзюк Т.В. Особенности клеточного состава крови серых тюленей (Halichoerus grypus) разного возраста // Зоол. журн. - 2011. - № 9. - С. 1122Ц1126.
- Минзюк Т.В. Возрастные изменения бактерицидной активности зернистых лейкоцитов серых тюленей // Вестник ЮНЦ. - 2011. - Т. 7, № 4. - С. 70Ц73.
Публикации в других изданиях
- Кавцевич Н.Н., Минзюк Т.В. Некоторые морфологические и цитохимические особенности крови щенков серого тюленя // Биология: Теория, практика, эксперимент: Матер. междунар. науч. конф. В 2-х кн. Саранск: ООО "Бьюти", 2008. - Кн. 2. - С. 167Ц171.
- Минзюк Т.В. Активность организаторов ядрышка в лимфоидных клетках крови щенков серых тюленей в различные периоды онтогенеза // Материалы XXVI конференции молодых ученых Мурманского морского биологического института, проводимой в рамках Международного полярного года (г. Мурманск, май 2008 г.). - Мурманск, 2008. - С. 92Ц96.
- Кавцевич Н.Н., Минзюк Т.В. Активность миелопероксидазы в лейкоцитах серых тюленей // Морские биотехнические системы. Биологические и технические аспекты. Тез. докл. Междун. научно-практ. конф. Ростов-на-Дону. 2008. - С. 76Ц78.
- Минзюк Т.В. Применение компьютерной морфометрии для оценки состояния лимфоидной системы морских млекопитающих. Морские биотехнические системы. Биологические и технические аспекты. Тезисы докл. междун. науч.-практ. конф. (г. Ростов-на-Дону, 10-11 сентября 2008г.). - Ростов н/Д: Изд-во ЮН - РАН, 2008. - С. 110Ц113.
- Минзюк Т.В., Кавцевич Н.Н. Содержание больших гранулярных лимфоцитов у гренландских тюленей разного возраста // Современные проблемы и методы экологической физиологии и патологии млекопитающих, введенных в зоокультуру. Матер. 4 междун. симп. Петрозаводск, 2009. - С. 181Ц186.
- Кавцевич Н.Н., Минзюк Т.В. Особенности клеточного состава крови щенков серого тюленя (Halihoerus grypus Fabricius, 1791) // Морские млекопитающие Голарктики: Материалы 5-й Междун. конф., 14-18 окт. 2008, Одесса. - Одесса, 2008. - С. 251Ц254.
- Минзюк Т.В. Районы организаторов ядрышка в лимфоцитах крови щенков серых тюленей // Ломоносов - 2009: Международная конференция студентов, аспирантов и молодых ученых; секция "Биология"; 13-18 апреля 2009 г.; Москва, МГУ имени М.В. Ломоносова, биологический факультет: Тезисы докладов. - М., 2009. - С. 268.
- Минзюк Т.В. Большие гранулярные лимфоциты крови гренландских тюленей в разные возрастные периоды // Материалы XXVII конференции молодых ученых Мурманского морского биологического института (г. Мурманск, май 2009 г.). - Мурманск, 2009. - С. 133Ц138.
- Кавцевич Н.Н., Минзюк Т.В. Клеточный состав крови щенков серых тюленей // Современные проблемы и методы экологической физиологии и патологии млекопитающих, введенных в зоокультуру. Материалы Международного симпозиума (23-25 сентября 2009 г., Петрозаводск, Республика Карелия, Россия). - Петрозаводск, 2009. - С. 103Ц108.
- Минзюк Т.В., Кавцевич Н.Н. Содержание больших гранулярных лимфоцитов у гренландских тюленей разного возраста. Современные проблемы и методы экологической физиологии и патологии млекопитающих, введенных в зоокультуру. Материалы Международного симпозиума (23-25 сентября 2009 г., Петрозаводск, Республика Карелия, Россия). - Петрозаводск, 2009. - С. 181Ц186.
- Кавцевич Н.Н., Минзюк Т.В. Выявление миелопероксидазы в лейкоцитах серых тюленей // Пробл. изуч. и охр. животного мира на Севере. Матер. всеросс. конф. Сыктывкар, 2009. - С. 327Ц328.
- Минзюк Т.В., Кавцевич Н.Н. Морфометрия районов организаторов ядрышка лимфоцитов серых тюленей // Пробл. изуч. и охр. животного мира на Севере. Матер. всеросс. конф. Сыктывкар, 2009. - С. 332Ц333.
- Минзюк Т.В., Кавцевич Н.Н. Клеточный состав крови щенков тюленя хохлача // Природа морской Арктики: современные вызовы и роль науки: Тез. докл. Междунар. науч. конф. (г.Мурманск, 10-12 марта 2010г.). - Апатиты: Изд-во Кольского научного центра РАН, 2010. - С.152Ц153.
- Минзюк Т.В., Кавцевич Н.Н. Морфологические особенности лейкоцитов щенков тюленя-хохлача // Соврем. научно-практ. достижения в ветеринарии. Сб. статей Междунар. конф. 15-16 апреля 2010г. - Киров, 2010. - С. 122Ц124.
- Минзюк Т.В. Лейкоцитарные индексы в оценке адаптационных реакций серых тюленей в постнатальном периоде развития // Материалы XXVIII конференции молодых ученых Мурманского морского биологического института, посвященной 100-летию со дня рождения М.М. Камшилова "Гидробиологические и экосистемные исследования морей европейского Севера" (г. Мурманск, май 2010 г.). - Мурманск: ММБИ КН - РАН, 2010. - С. 149Ц154.
- Минзюк Т.В., Кавцевич Н.Н. Клеточный состав крови щенков тюленя хохлача (Cystophoracristata Erxleben, 1777) // Морские млекопитающие Голарктики. Сб. научн. тр. по материалам VI междунар. конф. Калининград, 11-15 октября 2010г. - Калининград, 2010. - С. 397Ц401.
- Минзюк Т.В. Содержание катионных бактерицидных белков в гранулоцитах серых тюленей // Материалы XXIX конференции молодых ученых Мурманского морского биологического института, посвященной 140-летию со дня рождения Г.А. Клюге "Морские исследования экосистем Европейской Арктики" (г. Мурманск, май 2011г.). - Мурманск: ММБИ КН - РАН, 2011. - С. 147Ц150.
- Минзюк Т.В. Некоторые показатели естественной резистентности тюленей в постнатальном онтогенезе // Изучение и освоение морских и наземных экосистем в условиях арктического и аридного климата: материалы Международной научной конференции (6-11 июня 2011г., Ростов-на-Дону). - Ростов-на-Дону: ЮН - РАН, 2011. - С. 295Ц297.
24. Пат. 113178 Российская Федерация, МПК В04В 7/00 (2006.01), G01N 33/00 (2006.01). Устройство для изготовления препаратов биологических клеток на покровных стеклах центрифугированием / Ерохина И.А., Кавцевич Н.Н., Минзюк Т.В., заявитель и патентообладатель Учреждение Российской академии наук Мурманский морской биологический институт - № 2011141973/05. заяв. 17.10.2011, опубл. 10.02.2012, Бюл. № 4. - 2 с.
Список использованных сокращений
АОС - антиоксидантная система,
AgЯОР - окрашенный серебром ядрышкообразующий район,
БГЛ - большие гранулярные лимфоциты,
КБ - катионный белок,
КБ+лейкоцит - лейкоцит, содержащий катионный белок,
КБ+гранулы - окрашенные гранулы катионного белка,
МПО - миелопероксидаза,
СЦК - средний цитохимический коэффициент.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]