
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Род Trapa L. (Trapaceae Dumort.) в Приморском крае (особенности биологии, видовое разнообразие)
Автореферат кандидатской диссертации
На правах рукописи
БЕРЕСТЕНКО
Екатерина Николаевна
РОД TRAPA L. (TRAPACEAE DUMORT.) В ПРИМОРСКОМ КРАЕ
(ОСОБЕННОСТИ БИОЛОГИИ, ВИДОВОЕ РАЗНООБРАЗИЕ)
03.02.01 - Ботаника
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Владивосток - 2012
![]() Работа выполнена в лаборатории интродукции древесных растений Федерального государственного бюджетного учреждения Ботанического сада-института Дальневосточного отделения Российской академии наук
Работа выполнена в лаборатории интродукции древесных растений Федерального государственного бюджетного учреждения Ботанического сада-института Дальневосточного отделения Российской академии наук
Научный руководитель: аа кандидат биологических наук
Пшенникова Людмила Михайловна
Официальные оппоненты: аа Кожевников Андрей Евгеньевич
доктор биологических наук, старший научный сотрудник, заведующий лабораторией Биолого-почвенного института ДВО РАН
а Чубарь Елена Алексеевна
кандидат биологических наук, научный сотрудник Дальневосточного морского биосферного государственного заповедника ДВО РАН
Ведущая организация: аа Федеральное государственное бюджетное учреждение науки Институт водных и экологических проблем ДВО РАН, г. Хабаровск
Защита диссертации состоится л17 мая 2012 г. в 10-00 часов на заседании диссертационного совета Д 005.003.02 на базе Биолого-почвенного института ДВО РАН по адресу: 690022, г. Владивосток, проспект 100 лет Владивостоку, 159, конференц-зал.
Факс: (423) 2-310-193
E-mail: ibss@eastnet.febras.ru
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан аапреля 2012 г.

Ученый секретарь
диссертационного совета
доктор биологических наука аВ.Ю. Баркалов
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Территория Приморского края на Дальнем Востоке России представляет особый интерес в ботаническом отношении. Многие представители растительного мира региона являются прямыми наследниками аркто-третичной флоры. Большое число древнейших видов растений имеет в своем составе водная и прибрежная флора Приморского края (Куренцова, 1968; Пшеннникова, 2005).
Одним из реликтов третичного времени является род водяной орех или ронгульник (Trapa L.) - однолетнее водное растение, относящееся к монотипному семейству Trapaceae Dumort. Сложности изучения водяного ореха определянютнся, прежде всего, большим разнообразием его форм, в связи с чем взгляды иснследователей на вопросы его систематики неоднозначны. Помимо этого, водянные орехи - ценные пищевые и лекарственные растения, имеющие большое практическое применение в странах Восточной и Южной Азии (Индия, Китай, Япония). Также они могут служить прекрасным объектом для декоративного оформления пользующихся популярностью искусственных водоемов.
Несмотря на то, что представители рода Trapa широко распространены на юге российского Дальнего Востока, до сих пор нет единого мнения о таксономическом составе рода. Разные авторы для этой территории указывают от 3 (Ворошилов, 1982) до 8 (Цвелев, 1995) и даже 10 (Васильев, 1949) видов водяного ореха. В последние годы из Приморского края Л.М. Пшенниковой (2006, 2007) описано еще 3 новых вида. Причина разных мнений по видовому разнообразию кроется в различном подходе авторов к объему рода. В свою очередь различия в понимании объема рода неизбежно связаны, с одной стороны, с высоким уровнем полиморфизма, прежде всего, на уровне морфометрических показателей плодов, которым исследователи уделяют первоочередное внимание, и, с другой стороны, с явной нехваткой сведений об особенностях строения вегетативных и генеративных органов.
Противоречивы данные, касающиеся биологии цветения, опыления, хранения семян. Так, одни авторы (Михайлова, 1940) указывают на то, что цветки водяного ореха опыляются насекомыми, другие (Ram, 1956; Васильев, 1960) на возможность самоопыления. Неоднозначны также имеющиеся сведения относительно хранения семян. По одним данным семена водяного ореха при продолжительном пребывании на воздухе теряют всхожесть (Соловей, 1954; Пашкевич, Юдин, 1978), по другим - плоды водяного ореха жизнестойки и в иле сонхраняются в течение 50 лет, не прорастая при неблагоприятных условиях (Шлотгауэр, Мельникова, 1990). Открытым остается вопрос, касающийся продолжительности жизни водяного ореха. Считается, что это однолетнее растение, но есть указания на то, что водяной орех может быть и многолетним (Мурдахаев, 1975). В.Н. Васильев (1960) называет род Trapa потенциально многолетним, считая, что в умеренных широтах происходит насильственное окончание цветения с наступающими холодами, что делает это растение фактически однолетним, хотя биологически оно является многолетним.
В связи с этим, изучение полиморфизма и биологических особенностей водноореховых является актуальной задачей как для сохранении биоразнообразия, так и для повышения интереса к использованию этих растений в условиях культуры.
Цель работы. Изучить биологические особенности представителей семейства Trapaceae Приморского края, разработать рекомендации по их размнонжению в культуре и сформировать предпосылки для решения проблемы их систематики.
Задачи:
- Уточнить видовой состав и распространение водноореховых на территории Приморского края.
- Выявить внутривидовую изменчивость видов Trapa на уровне морфометрических признаков плодов.
- Оценить возможность формальной классификации (с использованинем современных методов кластерного анализа и распознавания образов) видов Trapa по морфометрическим показателям их плодов.
- Провести анализ анатомического строения стебля, черешка и плодоножки в связи с вопросами систематики.
- Изучить биологию цветения и плодоношения водяных орехов в условиях культуры.
- Исследовать влияние низких температур хранения плодов водяных орехов на всхожесть семян.
Научная новизна. аОбнаружены новые места произрастания T. incisa, T. maximowiczii, T. manshurica, T. japonica Fler., позволяющие расширить сведенния об их распространении в Приморском крае. Впервые изучен репродуктивный потенциал видов рода Trapa в условиях культуры на юге Приморского края. Установлено, что особенности анатомического строения плодоножки позволяют различать виды Trapa на уровне секций. Показана принципиальная возможность решения задачи видовой классификации представителей рода Traнpa по морфометрическим показателям их плодов. Выявлены таксономически значимые признаки плодов для дифференциации близких видов: T. khankensis Pshennikova и T. pseudoincisa Nakai, T. manshurica Fler. и T. kozhevnikoviorum Pshennikova, T. incisa Siebold et Zucc. и T. maximowiczii Korsh., T. nedoluzhkoi Pshennikova и Т. maximowiczii. Опыт решения данной задачи на базе современных методов распознавания образов имеет общее методологическое значение для проблемы систематики видов с выраженным полиморфизмом.
Практическая значимость. Разработаны рекомендации по хранению плодов и проращиванию семян водяных орехов, что наряду с данными по биологии цветения, плодоношения и семенной продуктивности этих растений может быть использовано для разведения их в культуре; на основе данных по морфометрии плодов составлен ключ для определения 8 видов водноореховых Приморского края. Данные по изучению анатомического строения плодоножки также могут быть использованы при рассмотрении вопросов систематики видов Trapa. В хонде полевых исследований получены сведения, уточняющие видовой состав водяных орехов и их распространение на территории Приморского края, что в свою очередь может послужить основой для разработки мер по охране водноореховых на указанной территории. Общее теоретическое и практическое значение для изучения видов рода Trapa имеют представленные в работе оценки статистических характеристик морфометрических показателей плодов.
Положения, выносимые на защиту:
- Традиционная схема линейной классификации позволяет успешно дифференцировать виды рода Trapa, произрастающие на территории Приморского края, по морфометрическим показателям их плодов.
- В условиях культуры на юге Приморского края водяные орехи являются однолетними растениями, но некоторые особенности вегетации, цветения и плодоношения указывают на то, что водноореховые обладают свойствами потенциально многолетних растений.
ичный вклад автора. Сбор материала и анализ данных, выбор методов, постановка экспериментов, получение результатов и их интерпретация осуществлены автором лично. В работах [1, 3] автор участвовал в постановке задачи, асборе данных и осуществлял их анализ.
Апробация работы. Результаты проведенных исследований были доложены и обсуждены на 62-ых Комаровских чтениях (Владивосток, 2008), на заседаниях Ученого совета Ботанического сада-института ДВО РАН (Владивосток, 2007, 2012), на заседании Приморского отделения Русского ботанического общества (Владивосток, 2012), на Конкурсе молодых ученых Зри в корень (Владивосток, 2012).
Публикации. По теме диссертации опубликовано 6 работ, в том числе две статьи в журналах, включенных в перечень ВАК.
Структура и объем работы. Диссертация состоит из введения, 7 глав, выводов, списка литературы и трех приложений. Работа изложена на 168 страницах машинописного текста, содержит 51 рисунок и 33 таблицы. Список литературы включает 170 источников, из которых 41 на иностранных языках.
Благодарности. Автор выражает искреннюю признательность своемуа научному руководителю к.б.н. Людмиле Михайловне Пшенниковой за предоставленную возможность заниматься изучением одной из интереснейших групп реликтовых водных растений Приморского края.
За помощь при проведении математических расчетов и статистической оценки результатов исследований автор благодарен к.ф.-м.н. Дмитрию Евгеньевичу Кислову, за ценные советы и рекомендации по улучшению работы д.б.н. Вячеславу Юрьевичу Баркалову, д.б.н. Светлане Борисовне Гончаровой, д.б.н. Ольге Викторовне Храпко, к.б.н. Роману Васильевичу Дудкину, к.б.н. Инне Михайловне Кокшеевой, к.б.н. Светлане Владимировне Нестеровой.
За веру и поддержку автор признателен д.б.н. Анне Витольдовне Беликович и д.б.н. Павлу Витальевичу Крестову, а также всем неравнодушным сотрудникам Ботанического сада-института ДВО РАН.
СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Ретроспективный анализ проблемы таксономии рода Trapa на Дальнем Востоке России
Глава носит вводный характер - в ней раскрываются проблемы систематики рода Trapa, существующие направления в ее решении. Приводится обзор основных работ отечественных и зарубежных исследователей, посвященных изучению этого вопроса (Флеров, 1925; Комаров, Клобукова-Алисова, 1932; Nakai, 1942; Васильев, 1949, 1965, 1973; Nakano, 1964; Ворошилов, 1966, 1982; Цвелев, 1993, 1995; Пшенникова, 2006, 2007). В частности указывается, что основные трудности в определении видов водяного ореха связаны с высоким уровнем полиморфизма, причину которого Н.Н. Цвелев (1993) связывает с гибридизацией между видами.
Также приводится сравнение видового состава водноореховых для флор сопредельных государств (Китай, Корея, Япония) и Приморского края (Nakai, 1942; Nakano, 1964; Hong, 1986; Kadono, 1994; Clavis PlantarumЕ, 1995; Li, аZheng, 1996; FloraЕ, 2000; Park C.-W., 2007).
Глава 2. Материалы и методы исследований
 2.1. Объекты и материал исследований
2.1. Объекты и материал исследований
В качестве объектов исследований в работе рассматриваются виды рода Trapa Приморского края, относящиеся к двум секциям. Секция Prototrapa Tzvel. представлена видами T. incisa, T. maximowiczii, T. nedoluzhkoi; секцияTrapaTzvel. видамиT. japonica, T. khankensis, T. kozhevnikoviorum, T. manshurica,а T. pseudoincisa.
![]() Материал для изучения внутривидовой изменчивости, особенностей хранения плодов, для анатомических исследований и проведения наблюдений за биологией цветения и плодоношения был получена в ходеа полевых работ в Дальнереченском, Кировском, Пожарском, Спасском, Уссурийском, Ханкайском и Хасанском районах Приморского края (рис. 1) в августе-сентябре 2005 и 2008 гг., а также в условиях культуры. Помимо этого, материалом для работы послужили фондовые коллекции гербариев Ботанического института им. В.Л. Комарова РАН (LE, БИН, г. Санкт-Петербург), Главного Ботанического сада им. Н.В. Цицина РАН (MHA, ГБС, г. Москва) и Биолого-почвенного института ДВО РАН (VLA, БПИ, г. Владивосток).
Материал для изучения внутривидовой изменчивости, особенностей хранения плодов, для анатомических исследований и проведения наблюдений за биологией цветения и плодоношения был получена в ходеа полевых работ в Дальнереченском, Кировском, Пожарском, Спасском, Уссурийском, Ханкайском и Хасанском районах Приморского края (рис. 1) в августе-сентябре 2005 и 2008 гг., а также в условиях культуры. Помимо этого, материалом для работы послужили фондовые коллекции гербариев Ботанического института им. В.Л. Комарова РАН (LE, БИН, г. Санкт-Петербург), Главного Ботанического сада им. Н.В. Цицина РАН (MHA, ГБС, г. Москва) и Биолого-почвенного института ДВО РАН (VLA, БПИ, г. Владивосток).
2.2. Методы исследований и статистической обработки данных
Согласно данным экспедиционных исследований, а также благодаря анализу гербарного материала были составлены карты распространения для каждого вида Trapa на территории Приморского края. Местонахождения видов наносились на карту с использованием точечного метода.
Изучение биологии цветения и плодоношения осуществлялось в условиях культуры. Наблюдения за цветением проводили с учетом рекомендаций А.Н. Пономарева (1960). Выполнялись наблюдения за цветением одного цветка, розетки, всего растения.
Расчет потенциальной и реальной семенной продуктивности, а также процента семенификации проводили на основе разработок И.В. Вайнагий (1973, 1974), Т.А. Работнова (1960, 1969). Под потенциальной семенной продуктивностью понимается количество семяпочек на особь или генеративный побег. Реальная семенная продуктивность - количество сформировавшихся семян. Процентное соотношение между этими показателями - процент семенификации, отражающий характер взаимоотношений организма с условиями их обитания. Плоды хранились в течение 6 месяцев (с октября по апрель) при пониженных температурах воды (+2С и +5С) в полной темноте. Для каждого вида в эксперименте было заложено по 50 штук плодов в трехкратной повторности. Оценка значимости выводов выполнялась на основе статистики ?2.
Анатомические исследования проводились на временных препаратах с учетом рекомендаций А.А. Никитина, И.А. Панковой (1982), Н.М. Державиной (2003). Рассматривали строение стеблей, листовых черешков, а также плодоножек 8 видов водноореховых. Материал для изучения был собран в конце периода вегетации и предварительно фиксировался в 75%-ном этаноле. Изготовление препаратов осуществлялось с использованием замораживающего микротома.
Внутривидовая изменчивость изучалась с использованием разработок С.А. Мамаева (1973) и Ю.А. Филипченко (1978) на уровне структурных признаков плодов. Для этого определялись их морфометрические параметры, для каждого из которых рассчитывались минимальное и максимальное значения, среднее арифметическое, среднеквадратическое отклонение (СКО) и коэффициент вариации. Доверительные интервалы для статистических характеристик рассчитывались в соответствии со стандартными схемами параметрической статистики (Кобзарь, 2006), либо на основе технологии имитации статистического выбора (Эфрон, 1988). Определялись наиболее изменчивые и константные морфометрические признаки.
Проверка соответствия выборочных данных нормальному закону распределения выполнялась с привлечением критерия Шапиро-Уилка и критериев согласия, использующих коэффициенты асимметрии и эксцесса (Кобзарь, 2006).
В качестве среды для выполнения расчетов и статистической обработки данных в работе используются интегрированные в единую систему посредством языка программирования Python ( свободно распространяемые библиотеки научных вычислений SciPy ( NumPy ( Общий однофакторный дисперсионный анализ (множественное сравнение) осуществлялся (на базе пакета SciPy) по следующей схеме. Предварительно устанавливался факт соответствия сравниваемых выборок нормальному распределению (тест Шапиро-Уилка с 5 % уровнем значимости), далее, если нормальность подтверждалась, устанавливался факт равенства дисперсий (критерий Бартлетта). В случае если равенство дисперсий подтверждалось, использовалась стандартная процедура однофакторного дисперсионного анализа Фишера. В противном случае (если выборки нормальны, а дисперсии неравны) - осуществлялось парное сравнение выборок с помощью критерия Уэлча (Кобзарь, 2006). В результате всех парных сравнений определялось минимальное критическое значение вероятности (p-value). Если гипотеза о нормальности отвергалась на начальном этапе, то применялся непараметрический тест Крускала-Уоллиса, которым устанавливался факт совпадения или различия средних значений для участвующих в анализе выборок.
Верификация результатов кластеризации выполнялась на двух уровнях: с привлечением внешней информации - на основе модифицированного индекса Рэнда (Gordon, 1999) (значение индекса Рэнда изменяется от 0 до 1; при нулевом значении наблюдается полное несоответствие полученной кластерной структуры с исходной (видовой), в случае его единичного значения - констатируется их полное соответствие), а также без нее - с использованием технологии имитации статистического выбора (Эфрон, 1988). Суть этой технологии заключается в многократном (в условиях проводимых ниже экспериментов 500-кратном) выполнении алгоритма кластеризации над случайными подвыборками (в экспериментах использовалось 80 %а данных от исходной совокупности), сформированными из классифицируемой совокупности. Далее, осуществляя анализ устойчивости парных появлений объектов ва одних и тех же кластерах и вводя соответствующие меры, позволяющиеа оценить эту устойчивость, оказывается возможным подобрать наилучшую классифицирующую ситуацию (алгоритм, число кластеров), приводящую к наиболее устойчивой кластерной структуре. Подробно даннаяа технология "устойчивого" кластерного анализа (consensus clustering) изложена в работах (Monti et.al., 2003; Simpson, 2010).
Для оценки точности решения задачи при использовании алгоритмов классификации по прецедентам рассматривались матрицы ошибок, элементы которых оценивались на основе схемы скользящего экзамена (Айвазян, 1989).
2.3. Структура базы данных
Приводится перечень измеряемых морфометрических показателей плодов водяного ореха, используемых в дальнейших расчетах. Рассматривается структура базы данных, включающая морфометрические описания видов, произрастающих на территории Приморского края: T. incisa (298 описаний), T. japonica (299 описаний), T. khankensis (100 описаний), T. kozhevnikoviorum (100 описаний), T. manshurica (316 описаний), T. maximowiczii (202 описания), T. nedoluzhkoi(22а описания), T. pseudoincisa (115 описаний).
В электронной форме база данных представлена файлами csv-формата с учетом места произрастания вида.
Предварительная обработка файлов измерений включала их фильтрацию для исключения возможных ошибок (Кобзарь, 2006), а также рандомизацию - имитацию погрешностей инструментальных измерений.
Глава 3. Общая характеристика водноореховых
3.1. Общая характеристика рода Trapa L.
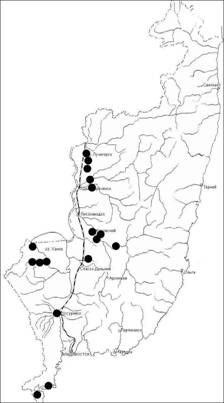
Приводится характеристика рода Trapa. Дается морфологическое описание растений, делается акцент на основных структурных элементах плодов на примере разных видов (рис. 2). Даются указания на те элементы плода, наличие или отсутствие, а также морфометрические характеристики которых могут иметь систематическое значение.
Рис. 2. Основные структурные элементы плодов водяных орехов: А - T. manshurica; Б - T. maximowiczii; В - T. japonica; Г - T. khankensis. 1 - коронка; 2 - шейка; 3 - верхние рога; 4 - пластрон; 5 - нижние рога; 6 - выросты на месте нижних рогов; 7 - граница пластрона; 8 - выросты на теле плода
Приводятся данные по общему распространению, центрах видообразования, экологии и охране рода Trapa(Васильев, Белавская, 1981; Красная книгаЕ, 1988; Редкие и исчезающие Е, 1981; Флеров, 1925).
3.2. Характеристика видов рода Trapa Приморского края
Приводится описание всех 8 видов, с которыми проводилась исследовательская работа, сопровождаемое фотографиями розетки листьев и плодов.
Глава 4. Распространение видов рода Trapa
на территории Приморского края и их охрана
Глава посвящена анализу распространения представителей рода Trapa на территории Приморского края. На основании собственных сборов и данных, полученных при изучении гербарных образцов, составлены карты распространения для всех видов (Пшенникова, Берестенко, 2006; Красная книгаЕ, 2008). Для видов T. incisa и T. manshurica указаны самые северные точки распространения на территории Приморского края (Пшенникова, Берестенко, 2006).
Виды водяных орехов характеризуются различной распространенностью на территории края. Так, одни виды охватывают практически всю территорию региона с севера на юг (T. maximowiczii), другие насчитывают всего по 1Ц2 известных места произрастания. Последние представлены тремя условно эндемичными видами - T. khankensis, T. kozhevnikoviorum и T. nedoluzhkoi).
Полученные данные могут послужить основой для проведения исследований, направленных на выявление степени угрозы исчезновения видов Trapa на территории Приморского края, что, в свою очередь, создает предпосылки для разработки мер по охране этих реликтовых растений.
Глава 5. Анализ внутривидовой изменчивости плодов рода Trapa
5.1. Статистические характеристики морфометрических признаков плодов
Установлено, что многие морфометрические показатели плодов видов Trapa обнаруживают существенное отклонение от нормального закона распределения. Построены оценки плотности (Парзена-Розенблатта) распределения некоторых морфометрических показателей (рис. 3). Обнаружена двумодовость распределений для отдельных морфометрических показателей. Двумодовость может указывать на наличие пары вполне устойчивых форм для данного вида, или на свойственный виду полиморфизм по данному морфометрическому признаку. Таким образом, делается вывод о необходимости использования методов непараметрической статистики при количественном анализе морфометрических характеристик плодов водяных орехов.
5.2. Характеристика внутривидовой изменчивости
Количественная оценка изменчивости морфометрических показателей плодов рода Trapa осуществляется с использованием коэффициента вариации (относительная изменчивость) и стандартного отклонения (абсолютная изменчивость).
Данные расчетов по наиболее изменчивым и наиболее константным признакам позволяют заключить следующее: 1) три наиболее изменчивых признака являются общими для всех видов; это признаки: длина, ширина и высота плода; 2) константные признаки представлены в более широком спектре и включают в себя группу признаков, описывающих шейку и коронку плода, верхние рога, нижние рога, а также расстояние от основания плода до основания нижних рогов; 3) виды секции Prototrapa характеризуются большей константностью морфометрических параметров по сравнению с видами секции Trapa.

Рис. 3. Оценки плотности распределения вероятностей (сплошная линия) для некоторых морфометрических показателей плодов T. manshurica (пунктир - соответствующее нормальное распределение): clen - длина коронки, lhl2 ? максимальная длина нижних рогов
5.3. Классификация плодов водяного ореха по морфометрическим параметрам
Для решения классификационных задач предварительно были сформированы группы видов по принципу принадлежности к той или иной секции и морфологической схожести плодов:
G1 - виды секции Prototrapa (T. incisa, T. nedoluzhkoi, T. maximowiczii);
G2 - виды секции Trapa с двурогими плодами (T. japonica, T. khankensis, T. pseudoincisa);
G3 - виды секции Trapa с четырехрогими плодами (T. kozhevnikoviorum, T. manshurica (Ильинские озера), T. manshurica (оз. Малое Мраморное)).
Дисперсионный анализ. Согласно критическим значениям вероятностей (p-value) значимое отличие средних для участвующих в каждой группе видов наблюдается практически по всем морфометрическим параметрам. Но к полученным результатам следует относиться с определенной долей осторожности, поскольку в реальной ситуации далеко не все эти признаки имеют решающее значение при дифференциации видов. Поэтому зачастую не последнюю роль играют экспертные представления об особенностях плодов водноореховых, которые в сочетании с полученными в результате расчетов данными способны обеспечить наиболее надежный процесс классификации.
Кластерный анализ. Сформированные в соответствии с несколькими алгоритмами кластера сопоставлялись с известным распределением видов по группам. В таблице 1 представлены результаты определения оптимального количества кластеров.
Таблица 1
Кластерный анализ для групп G1-G3
Группа |
Алгоритм кластеризации |
Индекс Рэнда |
||||
kmeans |
hclust |
pam |
agnes |
diana |
||
G1 |
2 |
3 (*) |
2 |
3 |
3 |
0.67 |
G2 |
2 |
2 |
3 |
3(*) |
2 |
1 |
G3 |
3(*) |
2 |
3 |
2 |
2 |
0.37 |
Примечание: * ? ситуация, соответствующая максимальному значению индекса Рэнда
Таким образом, согласно таблице 1 кластерная структура, получаемая при помощи метода кластеризации "agnes" с оценкой количества кластеров, в точности совпадает с видовой принадлежностью объектов ее образующих.
Дискриминантный анализ. Более конкретно оценить успешность дифференциации видов, составляющих группы G1-G3, можно исходя из результатов метода скользящего экзамена (Айвазян, 1989), который в данной работе применяется при классификации по прецедентам на базе линейных функций. Метод позволяет оценить долю ошибочной классификации объектов и, таким образом, дает представление о качестве алгоритма классификации.
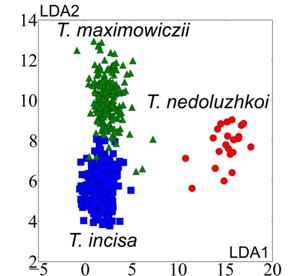
На рис. 4 в качестве примера приводятся данные обучающих выборок (двух исследуемых групп) в проекциях на главные дискриминантные оси.
 а
а
Рис. 4. Обучающие данные для групп G1, G2 в проекциях на главные дискриминантные оси
Ошибки классификации в отношении групп G1-G3 составили:
G1: T. incisa - 1.3 %, T. nedoluzhkoi - 0 %, T. maximowiczii - 2.5 %;
G2: каждый из видов идентифицируется абсолютно точно (ошибка 0 %);
G3: T. manshurica (Ильинские озера) - 34 %, T. manshurica (оз. Малое Мраморное) - 49 %, T. kozhevnikoviorum - 0%.
5.4. Ключ к определению видов Trapa Приморского края
Ключ представляет собой сформированное на основе обобщения экспертных представлений и результатов статистических исследований дерево решений, позволяющее определить вид водяного ореха по морфологическим параметрам его плода. Для возможности достоверного различия видов Trapa необходимо использовать сочетание метрических показателей плодов и генетически закрепленных особенностей их скульптуры. Признаки вегетативных органов и цветков, несомненно, имеют систематическое значение, но их применение на данный момент может носить лишь вспомогательный характер, поскольку в настоящее время не существует четких критериев, позволяющих достоверно определять виды с их помощью, а выявление таковых не входило в рамки настоящей работы. аВсе переходы по дереву решений, осуществляемые на основе количественных показателей, подтверждены в рамках традиционных моделей дисперсионного анализа.
1. Плоды сравнительно мелкие, 12Ц31 мм длиной по верхним рогам и 5Ц18 мм высотой. Поверхность тела плода, лишенного экзокарпия, гладкая, скульптура не развита. Плоды 4-рогие, рога отчетливо отделяются от тела плода ......ЕЕ..2
- Плоды среднего и крупного размера: 30Ц72 мм длиной и 11Ц32 мм высотой. Поверхность тела плода, лишенного экзокарпия, имеет обычно хорошо развитую скульптуру из бугров, вмятин и гребней. Плоды 2 или 4-рогие ...ЕЕЕ.Е4
2. Нижние рога отходят практически от основания плода: расстояние от основанния плода до основания нижних рогов 1Ц5 мм. Длина плода по верхним рогам больше ширины плода по нижним рога ...................................................................3
- Нижние рога отходят от середины тела плода: расстояние от основания плода до основания нижних рогов 5Ц9 мм. Длина плода по верхним рогам меньше или приблизительно равна ширине плода по нижним рогам..............ЕT. nedoluzhkoi
3. Шейка не расширяется в коронку. Листовые пластинки ромбические. Цветки розовые, мелкие (до 10 мм высотой). Плоды 12Ц22 мм длиной и 5Ц11,5 мм высотой ...............................................................................................................T. incisa
- Шейка с едва заметной или хорошо различимой коронкой. Листовые пластиннки с усеченным основанием. Цветки белые. Плоды 17Ц31 мм длиной и 8Ц17 мм высотой...............................................................................................T. maximowiczii
4. Плоды двурогие. Нижние рога отсутствуют, либо представлены короткими тупыми выростами, лишенными шипиков..............................................................5
- Плоды четырехрогие. Нижние рога хорошо развиты, как правило, несколько меньше верхних, имеют шипики ЕЕЕЕЕЕЕЕЕЕЕЕ.ЕЕЕЕЕЕЕ7
5. Плоды разнообразны по форме и величине. Шейка имеется, высотой 1Ц5 мм. Коронка округлой, вытянутой или лопастной формы, 3Ц21 мм длиной и 2,5Ц20 мм шириной................................................................................................................6
- Плоды веретеновидной или близкой к ней формы, за счет постепенно сужаюнщихся рогов, направленных горизонтально в стороны или несколько приподнятых. Шейка отсутствует, коронка лежит на теле плода или же незначительно над ним приподнимается. Длина и ширина коронки 3Ц7 мм ............T. khankensis
6. Рога тонкие, направлены косо вверх, высотой 1,5Ц3 мм. Шейка 2Ц4 мм толщиной, наверху расширяется в округлую коронку диаметром до 5 мм. Скульптура плода слабо развита, включая выросты на месте нижних рогов. Плоды 25Ц47 мм длиной и 10,5Ц20 мм высотой ЕЕЕЕЕЕЕЕЕ...ЕЕ.T. pseudoincisa
- Рога утолщенные, высотой 3Ц20 мм. Шейка 2Ц12,5 мм толщиной. Коронка 4,5Ц21 мм длиной и 4Ц20 мм шириной, округлой, квадратной, лопастной формы. Скульптура плода, как правило, хорошо развита. Выросты на месте нижних рогов могут достигать в длину 10 мм. Плоды 33Ц72 мм длиной и 12Ц32 мм высотой...........................................................................................................T. japonica
7. Шейка имеется. Коронка хорошо развита, лопастная, длина и ширина ее 5Ц14 мм. Верхние рога часто загнуты кверху ЕЕЕЕЕЕЕ..Е.Е.....T. manshurica
- Шейка отсутствует. Коронка прямоугольная, лежит на теле плода, длина и ширина ее 5Ц11 мм. Верхние рога покатые, горизонтально направленные Е.ЕЕЕЕЕЕЕЕЕЕЕЕЕЕ.........................................T. kozhevnikoviorum
Глава 6. Особенности анатомического строения водноореховых
в связи с вопросами систематики
Изучению анатомического строения различных органов водяного ореха посвящен ряд работ иностранных и отечественных авторов (Васильев, 1950, 1960, 1978; Лесков, 2006; Лотова, 2001; Тимонин, 1984, 1995; Couillault, 1972).
Анализируя литературу по данному вопросу, мы пришли к выводу, что как отечественные, так и зарубежные исследователи наибольший интерес проявляют к изучению анатомии семян и эмбриологии (Ghosh (Ram), 1954, 1956; Trela-Sawicka, 1978; Колесова (Титова), 1996, 2000; Titova et al., 1997; Титова, Захарова, 2000). Исследований, посвященных изучению вегетативных органов, меньше. Анатомические работы, проводимые с целью установления возможных систематических признаков, нам вовсе не встречались.
Принимая во внимание значительный полиморфизм видов Trapa и связанные с ним сложности при определении видов, в данной работе мы приводим результаты изучения анатомического строения стебля, черешка и плодоножки 8 видов водноореховых Приморского края для выявления возможных систематических отличий.
6.1 Анатомическое строение стебля
Приводится описание анатомического строения междоузлий. Междоузлия характеризуются первичным строением, что подтверждает литературные данные (Тимонин, 1995; Couillault, 1972). Результатом исследования является вывод о нецелесообразности использования особенностей строения стебля для решения задач систематики в связи с общностью его строения для всех 8 видов.
6.2 Анатомическое строение черешка
Приводится описание анатомического строения черешков. По результатам этого анализа делается вывод, что черешки разных видов водноореховых, как и стебли, имеют однотипное строение, и для решения задач систематики их использовать нецелесообразно.
6.3 Анатомическое строение плодоножки
Изучение анатомического строения зрелых плодоножек показало, что оно также может быть охарактеризовано по общей схеме. Вместе с тем, если говорить о значении строения плодоножки для систематики рода, то в данном случае основное внимание необходимо уделять ее срединной части. Так, представители секции Prototrapa с самыми мелкими плодами среди всех изученных водноореховых характеризуются отсутствием аэренхимы в срединной части плодоножек. Видам секции Trapa свойственно наличие воздухоносных полостей (рис. 5). Таким образом, особенности анатомического строения плодоножки позволяют различать виды Trapaна уровне секций.
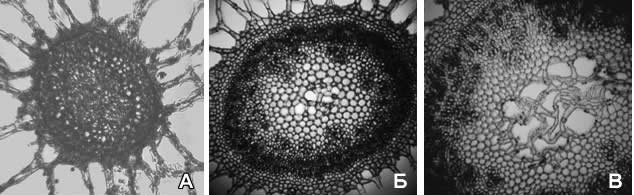
Рис. 5. Анатомическое строение срединной части плодоножки:
Секция Prototrapa: А - T. nedoluzhkoi
Секция Trapa: Б - T. kozhevnikoviorum; В - T. manshurica
Глава 7. Биологические особенности водноореховых
7.1. Биология цветения и плодоношения
В условиях культуры изучалось цветение и плодоношение 7 видов водяного ореха. В ходе наблюдений отмечали продолжительность цветения одного цветка, розетки, всего растения, продолжительность созревания плодов.
Период бутонизации характеризуется закладкой, формированием и ростом цветочных почек в пазухах листьев. Цветки водяных орехов одиночные, обоеполые, актиноморфные.
Раскрытие цветков водноореховых не имеет суточной приуроченности и может происходить как в утреннее, так и в послеобеденное время. Независимо от вида Trapa цветение одного цветка занимает период не более суток. Максимальное количество цветочных почек на одной розетке (генеративном побеге) может составлять 8Ц10 штук. Продолжительность цветения одной розетки, следовательно, зависит от количества цветочных почек на ней. То же самое можно сказать и в отношении целого растения, продолжительность цветения которого определяется количеством генеративных побегов. Поскольку на взрослом растении имеется, как правило, несколько генеративных побегов, появляющихся на протяжении всего периода вегетации и несущих в среднем по 4Ц6 цветочных почек, период цветения и плодоношения у всех водноореховых сильно растянут во времени. а
Рост плода начинается через 3Ц5 дней после отцветания. Период созревания плода с момента отцветания цветка до отделения от плодоножки составляет:
- от 30 дней для T. japonica, T. manshurica и T. kozhevnikoviorum;
- от 25 дней для T. khankensis, T. pseudoincisa и T. maximowiczii;
- от 20 дней для T. incisa.
Разница в продолжительности созревания может быть объяснена размерами плодов. У T. incisа они мелкие и, следовательно, для их развития требуется меньше пластического материала. T. japonica, T. manshurica и T. kozhevnikoviorum являются крупноплодными видами, а значит период развития плода заметно продолжительней.
Для трех видов в условиях культуры рассчитаны потенциальная семенная продуктивность (ПСП), реальная семенная продуктивность (РСП) и процент семенификации (табл. 2).
Таблица 2
Семенная продуктивность видов Trapa
Вид |
Число генерат. побегов |
ПСП, шт./ген.побег |
РСП, шт./ген.побег |
Процент семенификации |
T. khankensis |
28 |
7.36 0.65 |
2.82 0.3 |
38.32 |
T. japonica |
17 |
9.52 0.78 |
3.59 0.44 |
37.71 |
T. manshurica |
33 |
12.54 0.6 |
4.67 0.24 |
37.24 |
Показатель реальной семенной продуктивности оказывается меньше показателя потенциальной продуктивности в 3 раза. Причинами этого являются: 1) дегенерация одного из двух семязачатков (Титова, Захарова, 2000), 2) остановка в развитии некоторых цветочных почек, 3) короткий период вегетации, из-за чего не все плоды успевают созреть, 4) не у всех цветков происходит процесс оплодотворения.а
Интересным представляется явление повторного роста, наблюдаемое нами в условиях культуры почти у всех видов. Упоминание об этом явлении есть в публикации Ю.М. Мурдахаева (1975), посвященной опыту выращивания водяного ореха в условиях культуры в Узбекистане. В условиях культуры на юге Приморского края повторный рост начинается со второй половины августа при условии сохранения достаточно высоких температур воздуха и воды (не ниже 24С). Растения в этот период характеризуются интенсивным вегетативным приростом и закладкой большого количества цветочных почек. При этом цветочные почки располагаются не только в пазухах листьев, но и часто занимают верхушечное положение на вновь появляющихся побегах. Такие цветочные почки, как правило, не реализуют свой потенциал, а если же цветение происходит, то формирующиеся в результате него плоды не вызревают и характеризуются разного рода искаженной формой не свойственной данному виду.
Способность водяных орехов к повторному росту указывает на то, что эти растения имеют достаточно мощный вегетативный и генеративный потенциал, который в условиях нашего климата оказывается нереализованным. Это, в свою очередь, может служить подтверждением имеющихся в литературе фрагментарных данных и предположений о том, что водноореховые - многолетние растения. Но тот факт, что в условиях культуры по окончании периода вегетации все части растения отмирают, не оставляя зимующих органов, указывает на то, что в умеренном климате водяные орехи ведут себя, как однолетние растения. Подтверждением этому служат особенности их анатомического строения.
7.2 Влияние низких температур хранения на прорастание семян
Согласно классификации семян по характеристикам хранения, приводимой F.T. Bonner (1990), семена водноореховых следует отнести к группе лумеренных рекальцитрантов. Такие семена нельзя высушивать, но можно хранить в течение 3 - 5 лет при низких положительных температурах.
Приводятся результаты исследований влияния температурных условий хранения на всхожесть семян водяных орехов (табл. 3). Семена хранились при пониженных положительных температурах воды. Проращивались в лабораторных условиях при комнатной температуре воды после 5 - 6 месяцев хранения. Контролем послужили семена, хранившиеся в лабораторных условиях при комнатной температуре воды.
Таблица 3
Всхожесть семян видов Trapa при разных температурах хранения, %
(размеры выборок составляли по 50 объектов)
ВИД |
МЕСТО СБОРА |
+5С |
+2С |
c2 |
T.incisa |
Пожарское вдхр. |
80 5.8 |
83 3.3 |
0.55 |
окр. г. Владивостока |
76 8.8 |
86 3.3 |
5.01 |
|
T.japonica |
окр. г. Владивостока |
10 5.8 |
23 3.3 |
9.6 |
T.kozhevnikoviorum |
окр. Турьего Рога |
80 5.8 |
73 6.7 |
1.86 |
T.khankensis |
оз. Ханка, окр. с.Троицкое |
90 5.8 |
97 3.3 |
5.36 |
T.maximowiczii |
Пожарское вдхр. |
73 3.3 |
60 5.8 |
6 |
T.manshurica |
Ильинские озера |
60 10 |
67 8.8 |
1.44 |
оз.Ханка |
80 5.8 |
83 12 |
0.56 |
|
T.pseudoincisa |
Водоем вблизи оз. Утиного |
70 10 |
67 12 |
0.39 |
Примечание: c2 - статистика для оценки разницы в количестве проросших семян
Проведенные эксперименты позволили установить, что оптимальными условиями для хранения плодов большинства изучаемых видов Trapa являются пониженные положительные температуры воды, а именно +2?С и +5?С. При этом при температуре +2?С семена большинства видов демонстрируют более высокую всхожесть, что для T. incisa (Ботанический сад), T. japonica и T. khankensis является статистически подтвержденным (значения статистики c2 > 3,84 (Зайцев, 1984)). Контрольная группа семян характеризовалась отсутствием проросших образцов на протяжении всего периода хранения.
Таким образом, при пониженных положительных температурах (от +2?С до +5?С) плоды Trapa целесообразно хранить не более 5Ц6 месяцев. Причин этому две: во-первых, установлено, что длительный период хранения плодов водноореховых понижает всхожесть семян. Так, всхожесть семяна T. khankensis, хранившихся 6 месяцев, составилаа 97%, а семян, хранившихся 20 месяцев Ца 69%. Во-вторых, иногда семена начинают прорастать в условиях хранения, поэтому дальнейшее содержание их в данных условиях становится нецелесообразным, поскольку приводит к гибели проростков.
7.3. Рекомендации по хранению плодов и разведению водноореховых в культуре
- Период созревания плодов водяных орехов приходится на вторую половину августа и сентябрь. Собранные зрелые плоды необходимо хранить в воде при пониженных положительных температурах (от +2 до +5С). Перед хранением рекомендуется удалить экзокарпий плода, так как в нем неизбежно будут проходить процессы гниения, которые могут отрицательно сказаться на качестве семян. Особенно это важно для видов с некрупными плодами и тонкой наружной оболочкой, таких как T. incisa, T. khankensis, T. maximowiczii, T. pseudoincisa.
- Срок хранения плодов составляет 5Ц6 месяцев, таким образом, период окончания хранения приходится на мартЦапрель. Основная масса семян начинает прорастать в течение первой или второй недели после помещения плодов в лабораторные условия и характеризуется быстрым развитием проростков.
- Семена водяных орехов способны прорастать во время хранения, поэтому в течение всего этого периода необходимо периодически проверять семена на наличие среди них проросших. Если проросшие образцы обнаружены, то их хранение необходимо прервать и позволить проросткам развиваться в лабораторных условиях, так как в противном случае они погибают.
- Для посадки водяных орехов подходят водоемы глубиной до 80Ц100 см с илистым грунтом. Оптимальный срок высадки растений в открытый водоем - конец апреля и май, когда температура воды достигает 12Ц15С. Если высаживать растения позже, существует риск того, что плоды не успеют созреть. Также необходимо учитывать, что молодые неокрепшие и неукоренившиеся растения служат пищей для головастиков.
- Рекомендуемые виды для разведения в культуре: T. khankensis, T. incisa, T. manshurica, T. pseudoincisa. Семена этих видов характеризуются высокой жизнеспособностью, дружной всхожестью, а период созревания плодов укладывается в климатические условия юга Приморского края.
ВЫВОДЫ
- Подтверждено существование во флоре Приморского края 8 видов рода Trapa, относящихся к двум секциям.
- Константные признаки плодов Trapa представлены совокупностью характеристик, описывающих шейку и коронку плода, а также верхние и нижние рога. Данные признаки среди прочих являются таксономически значимыми для близких видов. Наибольшей константностью морфометрических параметров характеризуются плоды видов секции Prototrapa.
- Использование методов распознавания образов (в частности, алгоритма линейной классификации) позволяет успешно решить задачу морфометрической таксономии представителей рода Trapa Приморского края на основе анализа особенностей строения их плодов.
- Анатомическое строение плодоножки позволяет различать виды рода Trapa на уровне секций. Представители секции Trapa характеризуются различной степенью развитости воздухоносных полостей в срединной части плодоножки, а представители секции Prototrapa - отсутствием таковых.
- Растянутость сроков цветения и плодоношения, явление повторного роста и значительное количество незрелых плодов указывают на то, что водяные орехи являются потенциально многолетними растениями. Вместе с тем, отмирание подземных и надземных частей растений по окончанииа периода вегетации, а также особенности анатомического строения свидетельствуют о том, что водяные орехи в условиях сезонного умеренного климата Приморского края - однолетние растения.
- Продолжительность созревания плодов для крупноплодных видов водяных орехов составляет не менее 30 дней, для мелкоплодных - не менее 20 дней.
- Низкая реальная семенная продуктивность в условиях культуры обусловлена дегенерацией одного из двух семязачатков, остановкой в развитии некоторых цветочных почек и недоразвитием плодов.
- Оптимальные условия хранения плодов видов Trapa - пониженныеа положительные температуры воды от +2? С до +5? С, позволяющие сохранить высокую жизнеспособность семян в течение 5Ц6 месяцев.
- Разработаны рекомендации по хранению плодов и разведению водноореховых в условиях культуры на юге Приморского края.
Публикации по теме диссертации
Статьи, опубликованные в ведущих рецензируемых научных журналах:
- Пшенникова Л.М., Берестенко Е.Н. О находках редких водных растений на Дальнем Востоке России//Ботан. журн. - 2006. - Т. 91, № 12. - С. 1919Ц1921.
- Берестенко Е.Н. Жизнеспособность семян видов рода Trapa Приморского края при различных условиях хранения//Вестник ДВО РАН. - 2011. - №2. - С. 151Ц154.
Раздел в коллективной монографии:
- Пшенникова Л.М., Берестенко Е.Н. Водяной орех (выемчатолистный, японский, маньчжурский, Максимовича)//Красная книга Приморского края: Редкие и находящиесяа под угрозой исчезновения виды растений и грибов. - Владивосток: АВК Апельсин, 2008. - C. 219Ц224.а
Статья, опубликованная в электронном журнале:
- Берестенко Е.Н. Биология цветения и плодоношения видов Trapa в Ботаническом саду-институте ДВО РАН//Бюлл. БСИ ДВО РАН [Электронный ресурс] : науч. журн./Ботан. сад-институт ДВО РАН. - Владивосток, 2008. - Вып. 2. - С. 40-44. -
Работы, опубликованные в материалах международных научных конференций:
- Берестенко Е.Н. Trapa incisa Siebold et Zucc. в условиях культуры Ботанического сада-института ДВО РАН (г. Владивосток)//Матер. мiжнарод. наук. конф. Iнтродукцiя рослин на початку XXI столiття: досягнення i перспективи розвитку дослiджень. - Киiв: Фiтосоцiоцентр, 2005. - С. 64Ц66.
- Берестенко Е.Н. Биология цветения Trapa japonica Fler. в Ботаническом саду-институте ДВО РАН//Матер. международ. науч. конф. Биоморфологические исследования в современной ботанике. - Владивосток, 2007. - С. 69Ц71.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]