
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Разработка иммуностимулирующих комплексов на основе гликолипидов и тритерпеновых гликозидов из морских гидробионтов
Автореферат кандидатской диссертации
На правах рукописи
МАЗЕЙКА АНДРЕЙ НИКОЛАЕВИЧ
Разработка иммуностимулирующих комплексов на основе гликолипидов и ТРИТЕРПЕНОВЫХ ГЛИКОЗИДОВ из морских гидробионтов
03.01.06 - биотехнология (в том числе бионанотехнологии)
03.01.04 - биохимия
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
ВЛАДИВОСТОК
2012
Работа выполнена в Федеральном государственном автономном образовательном учреждении высшего профессионального образования Дальневосточный федеральный университет МОН РФ
Научные руководители адоктор биологических наук,
профессор
Санина Нина Михайловна
аа доктор биологических наук,
профессор
Костецкий Эдуард Яковлевич
Официальные оппонентыаа аБулгаков Виктор Павлович,
доктор биологических наук,
член-корреспондент РАН,
главный научный сотрудник
Биолого-почвенного института
ДВО РАНа
Ермак Ирина Михайловна,
доктор химических наук, а
старший научный сотрудник,
главный научный сотрудник
Тихоокеанского института
биоорганической химии
ДВО РАН а
Ведущая организация Федеральное государственное унитарное предприятиеа
Тихоокеанский научно-исследовательский
рыбохозяйственный центр, Владивосток
Защита состоится а15а маяаа 2012 г. ав а1000 часов на заседании диссертационного совета ДМ 005.003.04 на базе Биолого-почвенного института ДВО РАН по адресу: 690022 г. Владивосток, проспект 100 лет Владивостоку, 159.
факс 8(423) 2310-193
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан аа2 апреля 2012 г
Ученый секретарь
диссертационного совета
кандидат биологических наукаа аМузарок Т.И.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Разработка нового поколенияа вакцина связана с использованием высокоочищенных субъединичных антигенов, способныха повысить безопасность и протективность вакцин, получить высокоспецифичный иммунный ответ к бактериальным, вирусным патогенам и опухолевым клеткам. Серьёзным препятствием при создании субъединичных вакцин являетсяа недостаточная иммуногенность изолированных антигенов. Поэтому поиск эффективных адъювантов и новых способов презентации антигена иммунокомпетентным клеткам приобретает большую актуальность (Morein, Hu, 2000).
В последнее время пристальное внимание привлекают исследования мультимерной формы представления антигенов в виде иммуностимулирующих комплексов (ИСКОМ). ИСКОМ является стабильным комплексом холестерола (ХОЛ), фосфолипида и суммы сапонинов Qulajasaponaria Molina (QuilA). Иммунизация антигеном в составе ИСКОМ позволяет индуцировать реакции клеточного и гуморального иммунитета, направленные против вирусных и бактериальных антигенов (Kersten, Crommelin, 1995, 2000). Такой способ иммунизации в десятки раз усиливает образование специфических антител по сравнению с использованием убитых возбудителей или липосом (Lovgren, Morein, 1988).
Основной недостаток ИСКОМ - токсичность, обусловленная присутствием QuilA, разрушающего мембраны клеток (Kersten, Crommelin, 1995). Другой важный недостаток - отсутствие методики получения ИСКОМ, пригодной для коммерческого получения качественных препаратов комплекса с воспроизводимыми свойствами (Sun, Xie, 2009).а В связи с этим разработка нового эффективного и безопасного носителя антигенов, производство и применение которого было бы экономически выгодно,а представляет собой актуальную проблему.
Известно (Sjolander et al., 1997),а что различные носители и адъюванты не только усиливают иммунный ответ макроорганизма к антигену, но и обладают иммуномодулирующими свойствами. В связи с этим представляется важным использование природных соединений, подобных по физико-химическим свойствам компонентам классических ИСКОМ. Мы остановили свой выбор на изменении состава липидной компоненты ИСКОМ путем замены фосфолипидов, выделенных из яичного желтка, на гликоглицеролипиды из морских макрофитов, обладающих широким спектром биологической активности в отличие от иммунологически неактивных фосфолипидов. Так, известно, что моногалактозилдиацилглицерол (МГДГ) зеленых водорослей обладает противоопухолевой активностью (Morimoto et al., 1995), сульфохиновозилдиацилглицерол (СХДГ) ингибирует ДНК-полимеразу и обратную транскриптазу ВИЧ (Gustafson et al., 1998; Loya et al., 1998; Ohna et al., 1998).аа Кроме того, гликоглицеролипиды из морских макрофитов содержат до 80% полиненасыщенных жирных кислота (Saninaа et al., 2003, 2004), которые также иммуноактивны и могут модулировать иммуногенность комплекса. Следовательно,
_____________________________________
Использованные сокращения: ВЭЖХ - высокоэффективная жидкостная хроматография, ГА - гемагглютинин вируса гриппа, ДГДГ - дигалактозилдиацилглицерол, ИСКОМ - иммуностимулирующий комплекс, КД - кукумариозид (сумма моносульфатированных тритерпеновых гликозидов из С. japonica), КДА - кукумариозид А2-2, КДG Цкукумариозид G1, ГТА - голотоксин А1, МГДГ - моногалактозилдиацилглицерол, СХДГ - сульфохиновозилдиацилглицерол, ФРА - фрондозид А, ФХ - фосфатидилхолин, ХОЛ - холестерол, IL - интерлейкин, INF? - интерферон-гамма, QuilA - сумма тритерпеновых гликозидов Quillajasaponaria
использование тритерпеновых гликозидов голотурий, обладающих выраженной иммуностимулирующей активностью в сверхмалых концентрациях (Aminin et al., 2010),а вместо сапонина QuilA позволит повысить эффективность ИСКОМ, а способность тритерпеновых гликозидов голотурий полностью утрачивать токсические свойства в присутствии экзогенного холестерола (Попов,2002) позволит снизить токсичность ИСКОМ.
Цель и задачи исследования. Цель работы - разработка иммуностимулирующих комплексов на основе тритерпеновых гликозидов и гликолипидов из морских гидробионтов, оптимизация состава и метода получения адъювантного носителяа антигенов со стандартными свойствами, более низкой токсичностью и более высокой иммуногенностью, чем у классических ИСКОМ.
Для достижения данной цели были поставлены следующие задачи:
- Выбрать гликолипиды из морских макрофитов, пригодные для построения иммуностимулирующих комплексов.
- Выбрать сапонины (тритерпеновые гликозиды) из морских голотурий, пригодные для построения иммуностимулирующих комплексов.
- Исследовать токсичность (гемолитическую активность) выделенных гликолипидов и сапонинов.
- Установить соотношения между сапонинами, гликолипидами и холестеролом, обеспечивающие наиболее эффективное формирование модифицированных иммуностимулирующих комплексов.
- Оптимизировать способ получения иммуностимулирующих комплексов для достижения наиболее эффективного их формирования, последующего встраивания белкового антигена и получения препаратов со стандартными свойствами.
- Исследовать токсичность (гемолитическую активность) полученныха иммуностимулирующих комплексов и выбрать наиболее безопасный вариант.
- Выбрать оптимальный способ введения полученных иммуностимулирующих комплексов в организм млекопитающего, обеспечивающий наибольшую иммуностимулирующую активность.
Основные положения, выносимые на защиту.
1.Оптимальными для формирования модифицированного иммуностимулирующего комплекса являются МГДГ из морских макрофитов и тритерпеновый гликозид из Cucumariajaponica кукумариозид A2-2 (КДА).
2.Полученный комплекс, в отличие от ИСКОМ, является тубулярным иммуностимулирующим комплексом (ТИ-комплексом) и представляет собой тубулярные частицы с внешним диаметром около 16 нм, внутренним диаметром 6 нм и длинной около 500 нм.
3.Оптимальным для формирования ТИ-комплекса является весовое соотношение компонентов кукумариозид А2-2-холестерол-МГДГ 6:2:4.
4.ТИ-комплекс может служить носителем субъединичных антигенов бактериального и вирусного происхождения.
5.ТИ-комплекс обладает высокой адъювантной активностью в отношенииа тримера порина из Yersiniapseudotuberculosis.а Иммунизация субъединичным антигеном в составе ТИ-комплекса более эффективна, чем иммунизация в составе известных адъювантов: ИСКОМ и полного адъюванта Фрейнда, позволяет значительно повысить концентрацию специфических антител, не приводит к развитию побочных реакций.а
6.Оптимизированный ТИ-комплекс обладает низкой гемолитической активностью.
7. Иммунизацию необходимо проводить минимально разбавленным препаратома ТИ-комплекса.
Научная новизна и практическая значимость.
Впервые получен высокоэффективный липидный адъювантный комплекс с принципиально новым составом компонентова и суперструктурой. Впервые показано, что гликолипид МГДГ из морских макрофитов может эффективно замещать фосфолипиды в составе ИСКОМ, не изменяя их классической структуры. Впервые установлено, что тритерпеновый гликозид КДА из C. japonica способен заместить сапонины QuilA в иммуностимулирующих комплексах с преобразованием их везикулярной суперструктуры в тубулярную (тубулярные иммуностимулирующие комплексы или ТИ-комплексы). Впервые показано, что разработанный комплекс на основе МГДГ из морских макрофитов и КДА из голотурии C. japonicaобладает низкой токсичностью, высокой адъювантной активностью и способен инкорпорировать субъединичные антигены. Показана возможность использования ТИ-комплексов для создания вакцинных конструкций иа разработаны оптимальные условия их практического применения.
Выполненные исследования подкреплены двумя патентами Носитель антигенов (№2322259) и Иммуностимулирующий комплекс и его применение (№ 2010115549), которые могут быть использованы в практической медицине и ветеринарии.
Апробация работы и публикации.
Результаты работы были представлены на следующих конференциях: II региональной научной конференции Исследования в области физико-химической биологии и биотехнологии (Владивосток, 2006), X, XI и XIII международной молодежной школе-конференции по актуальным проблемам химии и биологии. (Владивосток, 2006, 2007 и 2010), The Third Asian Symposium on Plant Lipids (Yokohama, Japan, 2009), аIX региональной конференции студентов, аспирантов вузов и научных организаций Дальнего Востока России (Владивосток, 2010). 8th LIPIDOMICSа MEETING Membranes and Bioactive Lipids (Лион, Франция, 2011), 52nd International Conference on the Bioscience of Lipids, (Варшава, Польша 2011), The 4th Asian Symposium on Plant Lipids, (Hong Kong,а 2011).
Публикации. По теме диссертации опубликовано 20 работ, из них 8 статей в ведущих рецензируемых научных журналах.
Структура и объем работы.
Диссертация состоит из введения, 3 глав, выводов и списка литературы. Работа изложена на 154 страницах, иллюстрирована 40 рисунками и содержит 7 таблиц. Список литературы насчитывает 170 наименований.
Благодарности. Автор выражает благодарность Калинину В.И., Авилову С.А., Новиковой О.Д., Портнягиной О.Ю. (ТИБОХ ДВО РАН, Владивосток) за предоставленные вещества для проведения работы. Тимченко Н.Ф. (НИИ ЭМ СО РАН, Владивосток), Цыбульскому А.В., Веланскому П.В. (ГАОУ ВПО ДВФУ, Владивосток), Фомину Д.В. (ИБМ ДВО РАН, Владивосток) за помощь в проведении экспериментов.а Отдельно благодарю Попова А.М. (ТИБОХ ДВО РАН, Владивосток) за активное участие в обсуждении полученных результатов. Глубокую признательность и благодарность выражаю своим научным руководителям Костецкому Э.Я. и Саниной Н.М. за помощь и поддержку в работе.
Работа выполнена при финансовой поддержке Министерства образования и науки РФ (Госзадания 4.2007.2011 и 4.583.2011, Госконтракт П340), Правительства РФ (грант для государственной поддержки научных исследований, проводимых под руководством ведущих учёных в российских вузах, контракт 11.G34.31.0010) и CRDF (грант RUXO-003-VL-06-BP4M03).
МАТЕРИАЛЫ И МЕТОДЫ
5 видов морских макрофитов:а Ahnfeltiatobuchiensis,Sargassumpallidum, Laminariajaponica, Ulva fenestrata и Zosteramarina были использованы для выделения МГДГ.а Голотурия C. japonica была использована для выделения КДА. Испытания адъювантной и протективной активности полученных препаратов, проводили на мышах линий Balb\c, CBA и белых беспородных мышах массой около 20 г.
Сумма моносульфатированных гликозидов из С. japonica(Кукумариозид, КД), голотоксин А1 изApostichopusjaponicus(ГТА), фрондозид А (ФРА) из Cucumariafrondosaа и кукумариозид G1а (КДG) из Eupentactafraudatrixбыли выделены и любезно предоставлены в кристаллическом состоянии сотрудниками лаборатории химии морских природных соединений ТИБОХ ДВО РАН (зав. лаб. академик РАН, профессор, д.х.н. Стоник В.А.).
Порин из Yersiniapseudotuberculosis Pfeiffer (штамм 598, I серовар) был выделен и любезно предоставлен сотрудниками лаборатории молекулярных основ противобактериального иммунитета ТИБОХ ДВО РАН Новиковой О.Д. и Портнягиной О.Ю. При определении протективности ТИ-комплексов, содержащих порин, подопытных животных заражалиа Y. pseudotuberculosis (штамм 512, I серовар). Тесты на протективность были проведены на базе вивария НИИ эпидемиологии и микробиологии СО РАН, при любезной поддержке лаборатории молекулярных основ патогенности бактерий (зав. лаб. д.м.н. Тимченко Н.Ф.).
МГДГ получали путем хроматографии на силикагеле в системе хлороформ-ацетон 1:1, об/об. Полученную фракцию МГДГ очищали с помощью ТСХ в системе ацетон-хлороформ 1:1, об/об, иа ацетон-бензол-вода 91:30:6, об/об. Выделение ДГДГ и СХДГ проводили аналогично МГДГ, с использованием систем: ацетон-хлороформ 1:1, об/об, ацетон, ацетон-бензол-уксусная кислота-вода 200:30:3:10, об/об. Очистку проводили с помощью ТСХ, в системе ацетон-бензол-уксусная кислота-вода 200:30:3:10, об/об. Фосфатидилхолин выделяли из яичного желтка по методу (Singleton et al, 1965). Жирные кислоты липидов анализировали в форме метиловых эфиров методом газо-жидкостной хроматографии (Carreau, Dubacq, 1985).а
Кукумариозид А2-2выделялиа с помощьюа модифицированногоа нами метода, предложенного Авиловым С.А. и Калининым В. И с соавторами (Авилова и др., 1990). Стенку тела голотурии C. japonica экстрагировали 50%-ным этанолом,а из полученного экстракта получали кукумариозиды группы А2 путем последовательной хроматографии наа сорбенте Полихром-1 и силикагеле в системах хлороформ-этанол 3:1, об/об, хлороформ-этанол-вода 100:100:17, об/об. Препаративное выделение чистого КДА проводили на высокоэффективном жидкостном хроматографе SHIMADZU LC-8А с квадрупольным масс-спектрометром HPLC LCMS-2010EV в режиме отрицательных ионов, са использованием детекции по избранным ионам. Для выделения использовали обращенно-фазную хроматографию на колонкеа Sim-pack PREP-ODS 20 х 250 мм в градиенте концентраций метанола в воде. Концентрация метанола возрастала от 70% до 75% за 20 мин, затем от 75% до 90%а за 20 мин, и далее 90% метанола в течение 40 мин. Полученное вещество очищали с помощью препаративной высокоэффективной жидкостной хроматографии (ВЭЖХ) ана силикагеле (колонка Sim-pack PREP-sil 20 х 250 мм), в системе хлороформ-метанол-вода 73.2:22.95:3.8, об/об. Структуру полученного вещества идентифицировали методом 13С-ЯМР.
Для получения комплексов холестерол-тритерпеновый гликозид раствора холестеролаа в хлороформеа выпаривали насухо, к сухому остатку приливали водный раствор тритерпенового гликозида, фосфатно-солевой буфер и обрабатывали ультразвуком. Для получения ТИ-комплексов смешивали растворы МГДГ (или другого липида) и холестерола в хлороформе и выпаривали досуха. К сухому остатку прибавляли раствор КД, КДА или ГТА в воде и перемешивали инжектированием. Затем в смесь добавляли фосфатно-солевой буфер и подвергали ультразвуковой дезинтеграции. Для введения термоденатурированного порина в ТИ-комплекс, раствор белкаа одновременно с раствором тритерпенового гликозида добавляли к пленке, полученной после выпаривания растворов МГДГ и холестерола. Затем смесь обрабатывали ультразвуком.а Тримерную форму порина и гемагглютинин вируса гриппа в составе вакцины Influvac вводили в данный комплекс, добавляя раствор белка к липид-сапониновой суспензии немедленно после дезинтеграции. ИСКОМ получали по методу (Lovgren, Morein, 1988).
Исследование комплексов тритерпеновых гликозидов голотурий проводили на просвечивающем электронном микроскопе Libra120 (Zeiss, Германия) при ускоряющем напряжении 120 кВ. Использовали методы трансмиссионной электронной микроскопии с негативным контрастированием и окрашиванием гидрозолем железа (Гайер, 1973), а также метод трансмиссионной электронной микроскопии с энергетической фильтрацией (Energy Filtering Transmission Electron Microscopy - EFTEM) на неокрашенных образцах.
Для определения полноты включения антигеновпрепараты ТИ-комплексов подвергали ультрацентрифугированию при 45000 g в течение 30 мин, несвязанный белок определяли в надосадочной жидкости по методу Лоури (Lowry et al., 1951). Для определения агглютинирующей активности ТИ-комплекса с инкорпорированным в него гемагглютинином вакцины Influvac использовали реакцию прямой гемагглютинации с 0.5%-ной суспензией эритроцитов человека (0, Rh+). Для определения гемолитической активности выделенных тритерпеновых гликозидов, гликолипидов и липид-сапониновых комплексов в отношении эритроцитов человека использовали метод (Kersten et al., 1991). Реакцию торможения гемолизаа проводил по методике (Лиа и др., 2008).
При определении адъювантной активности полный адъювант Фрейнда вводили животным в объеме 200 мкл.а Термически денатурированную мономерную форму порина вводилиа в дозах 0.1,а 1 и 10 мкг, тример порина - 0.001, 0.1 и 1 мкг на мышь. ТИ-комплекс вводилиа в дозе 1 мкг КД/КДА на мышь. ТИ-комплекс КД-ХОЛ-МГДГ 3:2:6 вводили в объеме 200 мкл, тогда как ТИ-комплекс КДА-ХОЛ-МГДГ 6:2:4 вводили в объеме 10 мкл. Была использована двукратная схема иммунизации:а на 0-й и 7-й день. На 21-й день проводили либо забор сывороток крови, либо заражение животных.
Для определения количества специфических антитела и цитокинов в сыворотках крови использовали метод твердофазного иммуноферментного анализа.а При определении протективной активности ТИ-комплексов, животных внутрибрюшинно заражалиа 10ЛД50 Y. pseudotuberculosis.а В течение последующих 21 дня наблюдали динамику гибели животных.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Физико-химические свойства и ИСКОМ-формирующая способностьа гликоглицеролипидов из морских макрофитов
На первом этапе нами была исследована возможность модификации ИСКОМ путем замены фосфолипида различными по структуре гликоглицеролипидами (МГДГ, ДГДГ и СХДГ) из морского макрофита - бурой водоросли Laminariajaponica.
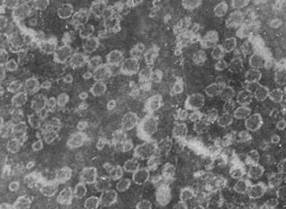
Указанные гликоглицеролипиды были использованы в качестве липидной компоненты для получения препаратов модифицированных ИСКОМ. Взятое нами соотношение холестерол-гликолипид-сапонины 1:1:4, по весу, как было показано в работе (Lovgren, Morein, 1988), являлось оптимальным для формирования везикул. Было установлено, что МГДГ и ДГДГ эффективно формировали ИСКОМ, тогда как в присутствии СХДГ везикулы не формировались. Включение МГДГ в ИСКОМ не изменяло классической морфологии везикул (Kersten, Krommelin, 1995). Модифицированные ИСКОМ обладали значительной гетерогенностью, размеры их варьировали от 16.5 нм до 33 нм. Коэффициент седиментации полученных частиц составлял 18-19 S. В результате включения ДГДГ в везикулы наблюдали два типа структур: ИСКОМ с диаметром 11-15 нм и спирали (рис. 1).
Вероятно, главной причиной наиболее эффективного включения МГДГ в ИСКОМ являлась его гексагональная суперструктура, способствовавшая формированию везикул с высокой кривизной поверхности, а также меньшая гидрофильность полярной части МГДГ по сравнению с другими гликоглицеролипидами (Israelashvili, 1992; Seddon, Templer, 1995).Основываясь на полученных данных, для дальнейшей модификации ИСКОМ использовали только один гликолипид - МГДГ.
 аа
аа 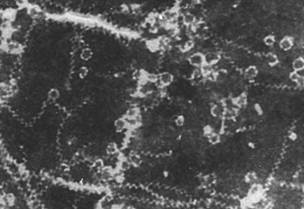
Б
Рис. 1. Электронная микрофотография везикул, полученных на основе МГДГ из L. japonica (увеличение ?189000) (А) и ДГДГ из L. japonica (увеличение ?231200) (Б)
Тритерпеновые гликозиды голотурий как компоненты иммуностимулирующих комплексова
В настоящей работе мы использовали 5 препаратов тритерпеновых гликозидов (рис. 2.), голотоксин А1,фрондозид А, кукумариозид G1, сумму моносульфатированных гликозидова из Cucumariajaponicaи кукумариозид А-2. Из 4 исследованных тритерпеновых гликозидов кукумариозид А2-2 из C. japonica по литературным данным обладал наибольшей иммуностимулирующей активностью, по этой причине данный тритерпеновый гликозид был выделен по модифицированной нами методике.
А
|
Б
|
В
|
Г
|




Рис. 2. Структурные формулы тритерпеновых гликозидов голотурий: (А) кукумариозид G1, (Б) кукумариозид А2-2, (В) фрондозид А, (Г) голотоксин А1, (Д)
В качестве основы нами был взят метод выделения тритерпеновых гликозидов из C. japonica, разработанныйаа Авиловым С.А. и Калининым В. И с соавторами а(Авилов и др., 1990). В данную методику нами была введена дополнительная стадия экстракции суммы тритерпеновых гликозидов, получаемой в результате хроматографии на Полихроме-1 системамиа хлороформ-этанол-вода 100:75:10 и 100:100:17, об/об. Также мы исключили многократную хроматографию на силикагеле и ограничились двукратным разделением. Вместо ВЭЖХ с рефрактометрической детекцией мы использовали тандемную ВЭЖХ-масс спектрометрию. Использование данного метода в режиме детекции по избранным ионам позволило выделять чистый кукумариозид А2-2 даже из фракций сложного состава. Внесенные изменения позволили сократить трудоемкость методики. Структура полученного кукумариозида А2-2 была подтвержденаа методом 13С-ЯМР-спектроскопии.а
аКомплекс тритерпеновый гликозид-холестерол как основа иммуностимулирующего комплекса
На следующем этапе исследований мы попытались заменить другой компонент ИСКОМ - сапонины QuilAа на тритерпеновые гликозиды голотурий с целью повышения иммуногенности и снижения токсичности иммуностимулирующих комплексов. Электронномикроскопическое исследование показало, что все изученные тритерпеновые гликозидыа оказались способны формировать надмолекулярные комплексы с холестеролом (рис. 3).
Так, кукумариозид А2-2, оказался способным формировать надмолекулярные комплексы с холестеролом, частицы которых имели четко выраженное тубулярное строение и несли гидрофильный канал в центре. На формирование частиц надмолекулярных комплексов данным тритерпеновым гликозидом большое влияние оказывало соотношение КДА-ХОЛ,а при возрастании молярной доли холестерола частицы надмолекулярных холестерол-сапониновых комплексов приобретали более выраженную тубулярную структуру (рис. 3 А, Б).
В отличие от чистого кукумариозида А2-2, сумма моносульфатированных тритерпеновых гликозидов (Кукумариозид, КД) из той же голотурии C. japonicaформировала надмолекулярные комплексы с холестеролом в водной среде только при большой его доле.
А
|
В
|
Д
|
Б
|
Г
|
Е
|


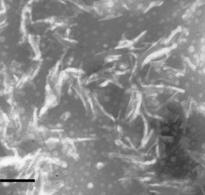
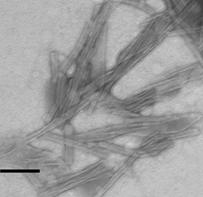
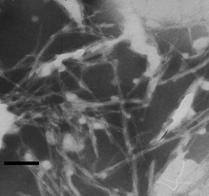
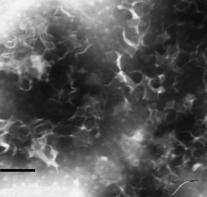
Рис. 3. Электронные микрофотографии структур, образованных системами с различным молярным соотношением компонентов: А - КДА-ХОЛа 1:1; Б - КДА-ХОЛ 1:9;. В - ГТА-ХОЛ 1:1; Г - ГТА-ХОЛ 1:2; Д - ФРА-ХОЛ 1:2; Е - КДG-ХОЛ 1:2. Масштабная линия соответствует 100 нм.а
Частицы, обнаруженные в системах ГТА-ХОЛ 1:1 и 1:2 обладали тубулярной структурой. В пределах обнаруженных частиц чередовались нитевидные и тубулярные участки (рис. 3 В, Г). Увеличение содержания холестерола в комплексе ГТА-ХОЛ 1:2 приводило к формированию более однородных по структуре частиц, однако при этом был обнаружен избыток холестерола в виде самостоятельной фазы (рис. 3 Б). Частицы надмолекулярных комплексов голотоксина А1 с холестеролом, в отличие от таковых кукумариозида А2-2, по мере увеличения доли холестерола в системе не формировали четко выраженных тубул.
Фрондозид А и кукумариозид G1 формировали надмолекулярные комплексы с холестеролом с нечетко выраженной структурой. По сравнению с кукумариозидом А2-2 и голотоксином А1 данные тритерпеновые гликозиды обладали худшей способностью к формированию надмолекулярных холестерол-сапониновых комплексов, тубулярного типа строения.
Полученные результаты показали, что кукумариозид А2-2, голотоксин А1 и фрондозид А формировали с холестеролом надмолекулярные комплексы тубулярного типа, отличающиеся от комплексов кукумариозида G1, не имевших тубулярного строения. В ряду КДАЦГТАЦФРА степень выраженности тубулярной структуры соответствующих комплексов тритерпеновый гликозид-холестерол убывала. В целом ГТА и КДG оказались не пригодными для модификации ИСКОМ. Таким образом, для дальнейшей разработки адъювантного носителя антигенов были выбраны кукумариозид А2-2 и голотоксин А1
Введение мембранных липидов в комплексы кукумариозид-холестерол и голотоксин А1-холестерол
В состав классических ИСКОМ, кроме тритерпеновых гликозидов и холестерола, входит также мембранообразующий липид: фосфатидилхолин (ФХ) или фосфатидилэтаноламин. Липид необходим как связующее звено в структуре ИСКОМ, а также нужен для встраивания субъединичных антигенов (Kersten et al., 1991).а Поэтому, мы исследовали формирование надмолекулярных комплексов тритерпеновый гликозид-холестерол в присутствие мембранообразующих липидов ФХ и МГДГ.
Для исследования комплексов с липидами вместо чистого кукумариозида А2-2 была использована сумма моносульфатированных тритерпеновых гликозидов из C. japonica - КД. В состав КД входят в равном соотношении кукумариозиды А2-2 и А4-2 и, в небольшом количестве, другие моносульфатированные гликозиды. КД, также как и КДА,обладает иммуностимулирующей активностью, но его получение экономически более выгодно.
А
|
Б
|
В
|
Г
|

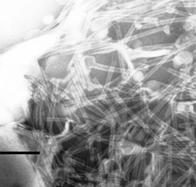
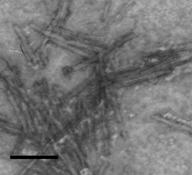

Рис. 4. Электронные микрофотографии структур, образованных системами тритерпеновый гликозид - холестерол с различными молярными соотношениями компонентов:а А - КД-ХОЛ-МГДГ 1:2:3; Б - КДЦХОЛ-ФХ 1:2:3; В - ГТА-ХОЛ-МГДГ 3:2:1; Г - ГТА-ХОЛ-ФХа 1:1:3. Масштабная линия - 100 нм.
аВо всех исследованных системах КД-ХОЛ-МГДГ нами были обнаружены тубулярные наночастицы, с четкими очертаниями и внутренним каналом диаметром 2-6 нм;а наружный диаметр обнаруженных частиц составлял 10-16 нм, а длинна - около 300 нм (рис. 4 А). Введение МГДГ в комплекс КДА-ХОЛ способствовало формированию тубулярных частиц постоянного внешнего и внутреннего диаметра с четкими краями при гораздо меньшем молярном соотношении гликозид-холестерол, чем в его отсутствии - 1:2 и 1:9 соответственно. Увеличение содержания МГДГ от 0.5 до 5 молярных частей в системе КД-ХОЛ-МГДГ с постоянным молярным соотношением КД-ХОЛ 2:1 не приводило к формированию самостоятельной липидной фазы. Замена МГДГ на ФХа приводила к формированию в системе КД-ХОЛ-ФХ 3:2:1 искаженных тубулярных частиц. В отличие от МГДГ, часть ФХ не входила в состав липид-сапониновых комплексов (рис. 4 Б).
Введение в систему ГТА-ХОЛ липидов резко улучшало формирование тубулярных частиц с четкими краями и внутренним каналом диаметром 3-5 нм, наружным диаметром 9-14 нм и длинной около 100 нм (рис. 4 В). Повышение молярной доли МГДГ в комплексе ГТА-ХОЛ-МГДГ от 1:2:1 до 1:2:3 приводило к проявлению частиц с постоянным внутренним и внешним диаметром с четкими краями. В системе, полученной в результате замены МГДГ на ФХ, наблюдались частицы с четко выраженной тубулярной структурой, диаметром внутреннего канала 2-4 нм и наружным диаметром 12-14 нм в отличие от системы ГТА-ХОЛ 1:1. Также наблюдался избыток ФХ в виде самостоятельной фазы (рис. 4 Г).
Из полученных данных видно, что присутствие липида МГДГ или ФХ было также необходимо для формирования надмолекулярных комплексов на основе тритерпеновых гликозидов голотурий, как и наличие холестерола. Введение в липид-сапониновый комплекс МГДГ представляется более предпочтительным, чем введение ФХ, так как позволяло получать гомогенные препараты липид-сапониновых комплексов, содержавших частицы только одного типа. На основании полученных данных о формировании тубулярных наночастиц для дальнейшей работы нами был выбран комплекс КД-ХОЛ-МГДГа с молярным соотношением компонентов 1:2:3, что соответствовало весовому соотношению 3:2:6. Этот комплекс в последствие был назван нами тубулярным иммуностимулирующим комплексом (ТИ-комплексом).
Структура частиц тубулярного иммуностимулирующего комплекса
 С целью установить локализацию углеводных цепей тритерпеновых гликозидов в тубулярных частицах было использовано позитивное окрашивание кислым гидрозолем окиси железа. Принцип окрашивания состоит в отложении электронноплотного вещества в местах локализации кислотных групп полисахаридов (сульфатных групп тритерпеновых гликозидов). На полученных микрофотографиях частицыа комплекса КД-ХОЛ-МГДГа 3:2:6, были окрашены снаружи, тогда как внутренний канал практически не был окрашен (рис. 5). Это позволило полагать, что углеводные цепи молекул кукумариозидов А2-2 и А4-2 экспонированы преимущественно на внешней стороне частиц. Такое расположение молекул сапонинов хорошо согласуется с предложенной моделью строения надмолекулярных комплексов гликоалкалоидов растений семейства пасленовые (Keukens et al., 1995), и позволило полагать, что данная модель в общих чертах справедлива и для разработанного нами носителя.
С целью установить локализацию углеводных цепей тритерпеновых гликозидов в тубулярных частицах было использовано позитивное окрашивание кислым гидрозолем окиси железа. Принцип окрашивания состоит в отложении электронноплотного вещества в местах локализации кислотных групп полисахаридов (сульфатных групп тритерпеновых гликозидов). На полученных микрофотографиях частицыа комплекса КД-ХОЛ-МГДГа 3:2:6, были окрашены снаружи, тогда как внутренний канал практически не был окрашен (рис. 5). Это позволило полагать, что углеводные цепи молекул кукумариозидов А2-2 и А4-2 экспонированы преимущественно на внешней стороне частиц. Такое расположение молекул сапонинов хорошо согласуется с предложенной моделью строения надмолекулярных комплексов гликоалкалоидов растений семейства пасленовые (Keukens et al., 1995), и позволило полагать, что данная модель в общих чертах справедлива и для разработанного нами носителя.
Рис. 5. Электронная микрофотография комплекса КД-ХОЛ-МГДГ с весовым соотношением компонентов 3:2:6, окрашенный гидрозолем окиси железа. Масштабная линия соответствует 100 нм.
Гемолитическая активность комплексов голотоксин А1Цхолестерол-липид и кукумариозид-холестерол-липид
Хорошо известно, что гемолитическая активность сапонинов QuilA сохраняется даже в составе ИСКОМ, несмотря на входящий в их состав холестерол. Гемолитичность ИСКОМ является главным препятствиема для их широкого практического применения. В связи с этим мы провели исследование гемолитической активности голотоксина А1, кукумариозида и их комплексов с холестеролом и мембранными липидами в сравнении с гемолитической активностью QuilA.
Голотоксин А1 и кукумариозид, по сравнению с растительными сапонинами QuilA, оказались более токсичным в 4 и 5 раз, соответственно. Полученные нами данные по нейтрализующему действию холестерола показали, что сапониныQuilA, полностью утрачивали гемолитическую активность в присутствии 2-х молекул холестерола на 1 молекулу сапонина. В случае голотоксина А1 100%-ное торможение гемолиза имело место только при соотношении холестерол-гликозид 8:1, а максимальный уровень подавления гемолитической активности кукумариозида при таком же соотношении холестерол-гликозид не превышал 50%. Отсутствие отмены гемолитической активности кукумариозида, возможно,а свидетельствовало в пользу существования дополнительных механизмов гемолитического действия данного вещества, не связанных со стеринами.
Для проверки этого предположения мы оценили гемолитическую активность тритерпеновых гликозидов голотурий в препаратах, содержавших кроме холестерола мембранообразующие липиды - МГДГ или ФХ.а Из полученных данных (табл. 1) видно, что, дополнительное введение в систему МГДГ или ФХ приводило к многократному увеличению коэффициента ингибирования гемолитической активности.а
Таблица 1
Гемолитическая активность кукумариозида и голотоксина А1 в составе липид-сапониновых комплексов
Комплекс |
Молярное соотношение |
Коэффициент ингибирования* |
К-ХОЛ |
1-2 |
2 |
КД-МГДГ |
1-3 |
1 |
КД-ФХ |
1-3 |
1 |
КД-ХОЛ-МГДГ |
1-2-3 |
8 |
КД-ХОЛ- ФХ |
1-2-3 |
8 |
ГТА-ХОЛ |
1-2 |
5.3 |
ГТА-МГДГ |
1-3 |
1.2 |
ГТА-ФХ |
1-3 |
1.5 |
ГТА-ХОЛ-МГДГ |
1-2-3 |
9.8 |
ГТА-ХОЛ- ФХ |
1-2-3 |
9.8 |
Коэффициент ингибирования рассчитывали из соотношения ЕС50 гликозидов в составе липидных комплексов к ЕС50 индивидуальных тритерпеновых гликозидов.
Полученные данные свидетельствовали о том, что в присутствии мембранообразующих липидов взаимодействие тритерпеновых гликозидов с холестеролом происходило иначе, чем в их отсутствие. Вероятно, ведущую роль в этом процессе играло взаимодействие агликона тритерпеновых гликозидов с холестеролом (Popov, 2003), тогда как наличие мембранообразующих липидов стабилизировало образующийся комплекс. В результатеа добавление в среду только холестерола оказывалось недостаточным для подавления гемолиза исследованными тритерпеновыми гликозидами. Надо полагать, что мембранообразующие липиды (МГДГ или ФХ), способствовали увеличению термодинамической устойчивости стерин-гликозидного комплекса за счет создания гидрофобной среды, в которойа усиливалось взаимодействие холестерола с агликоном гликозида.а
Вместе с тем присутствие мембранных полярных липидов не является необходимым условием для существования различных надмолекулярных стерин-сапониновых комплексов, устойчивых даже к действию детергентов (Keukensа et al., 1995). Ранее было показано (Попов, 2002; Popov, 2003) что высокоактивные тритерпеновые гликозиды голотурий, например голотурин А, эхинозид А, кукумариозид G1, обладали высоким аффинитетом к холестеролу как в липосомальных мембранах, так и в водной среде. Что касается кукумариозида, то его эффективное взаимодействие с холестеролом, очевидно, возможно только в присутствии липидов, то есть в гидрофобной среде.
Способы введения антигенов в тубулярный иммуностимулирующий комплекс
Для дальнейших экспериментов по разработке носителя субъединичных антигенов нами был выбран липид-сапониновый комплекс с молярным соотношением компонентов КД-ХОЛ-МГДГ 1:2:3, что соответствовало весовому соотношению 3:2:6. Мы исследовали возможность включения в данный комплекс модельных антигенов, в качестве которых использовали гемагглютинин вируса гриппа из вакцины Influvac и порообразующий белок (порин) из Y. pseudotuberculosis.
Гемагглютинин (ГА) вакцины Influvac - вводили в ТИ-комплекс так, что весовое соотношение компонентов КД-ХОЛ-МГДГ-ГА в итоге составляло 3:2:6:1.5 и 3:2:6:0.15. Для введения антигенов использовалось два различных способа: в первом случае - раствор антигенов прибавляли одновременно с раствором КД к пленке холестерола и МГДГ, а затем проводили ультразвуковую дезинтеграцию; во втором случае - раствор антигенов вводили в липид-сапониновую суспензию непосредственно после обработки ультразвуком.
В системе КД-ХОЛ-МГДГ-ГА с весовым соотношением компонентов 3:2:6:1.5 применение первого способа введения антигена в носитель приводило к эффективному встраиванию антигенов (72%), тогда как использование второго способа приводило к резкому падению эффективности включения антигена (21%). При меньшем количестве антигена в системе КД-ХОЛ-МГДГ-ГА 3:2:6:0.15 введение с применением обоих способов оказалось одинаково эффективно (54%). Таким образом, ультразвуковая дезинтеграция способствовала эффективному включению антигенов вакцины Influvac в ТИ-комплекс.
Оценка функциональной активности гемагглютинина вакцины Influvac включенного в состав разрабатываемого носителя показала, что при введенииа антигенов в носитель после дезинтеграции, полученный комплекс обладал более высокой агглютинирующей активности, чем, когда введение антигена производили до ультразвуковой обработки, несмотря на то, что полнота включения антигена при этом повышалась. Такое явление, вероятно, было связано с повреждением макромолекул гемагглютининаа ультразвуком. Полученные данные говорили о том, что наиболее подходящима способом введения гемагглютинина вакцины Influvac в комплекс КД-ХОЛ-МГДГ 3:2:6 являлось добавлениеа антигена к полученному комплексу непосредственно после обработки ультразвуком.
Другой модельный антиген - амфифильный порообразующий белок наружной мембраны возбудителя псевдотуберкулеза Y. pseudotuberculosis, с молекулярной массой около 36 кДа (порин), имел, в отличие от гемагглютинина, бактериальное происхождение. Порин в термически денатурированной мономерной форме вводили в ТИ-комплексы, так что весовое соотношение компонентов КД-ХОЛ-МГДГ-порин составляло 3:2:1:3, 3:2:6:3 и 3:2:10:3. Определение полноты инкорпорирования данного белка показало, что в комплекс КД-ХОЛ-МГДГ 3:2:1 включалось 92% антигена, тогда как в липид-сапониновые комплексы с весовым соотношением компонентов 3:2:6 и 3:2:10 включалось 88% и 90% белка соответственно.
 Было обнаружено, что введение гемагглютинина из вакцины Influvac приводило к появлению своеобразных "шапочек" на торцах типичных тубулярных частиц (рис. 6). В свою очередь, введение порина в комплекс не приводило к изменению его структуры, за исключением увеличения внешнего и внутреннего диаметра частиц, примерно, на 2 нм. Полученные данные позволяют утверждать, что липид-сапониновые комплексы КД-ХОЛ-МГДГ с весовым соотношением компонентов 6:2:1-10 могут быть использованы как носители субъединичных белковых антигенов бактериального и вирусного происхождения.
Было обнаружено, что введение гемагглютинина из вакцины Influvac приводило к появлению своеобразных "шапочек" на торцах типичных тубулярных частиц (рис. 6). В свою очередь, введение порина в комплекс не приводило к изменению его структуры, за исключением увеличения внешнего и внутреннего диаметра частиц, примерно, на 2 нм. Полученные данные позволяют утверждать, что липид-сапониновые комплексы КД-ХОЛ-МГДГ с весовым соотношением компонентов 6:2:1-10 могут быть использованы как носители субъединичных белковых антигенов бактериального и вирусного происхождения.
Рис. 6. Электронная микрофотография структур, образованных системой КД-ХОЛ-МГДГ-ГА с весовым соотношением компонентова 3:2:6:0.15.а Масштабнаяа линия - 100 нм.
Адъювантная активность тубулярного иммуностимулирующего комплекса кукумариозид-холестерол-моногалактозилдиацилглицерол 3:2:6
Для оценки адъювантной активности ТИ-комплексаа был использован комплекс КД-ХОЛ-МГДГ 3:2:6, несущий порин в термически денатурированной мономерной форме. Полученные данныеа (рис.7) продемонстрировали, что дозы порина 0.1, 1 и 10 мкг на мышь обеспечивали практически одинаковый уровень специфического гуморального иммунного ответа, тогда как иммунизация порином в этих же дозах, но в составе ТИ-комплексов обеспечивала более высокие показатели.аа При этом доза поринаа 0.1 мкг/мышь в составе ТИ-комплекса вызывала более высокий гуморальный иммунный ответ, чем введение чистого порина в дозе 1 мкг/мышь. Это позволяет говорить о способностиа ТИ-комплекса на порядок повышать эффективность иммунизации в сравнении с индивидуальным порином. Сравнение адъювантной активности ТИ-комплексов и известных носителей показало, что при внутрибрюшинном введении порина в дозе 10 мкг/мышь ТИ-комплекс обладал более высокой адъювантной активностью,а чем комплексы порина с полным адъювантом Фрейнда и ИСКОМ (рис.8).
Исследование изменения концентрации цитокинов при иммунизации порином выявило значительное угнетение интерферонового статуса (рис. 9), что свидетельствовало о негативных результатах иммунизации чистым порином. Иммунизация порином в составе ТИ-комплекса нейтрализовала супрессорный эффект порина в отношении интерферона-гамма (INF?), посколькуа сам по себе ТИ-комплекс вызывал значительное повышение уровня INF? в сыворотке крови животных. Вместе с тем иммунизация порином в составе ТИ-комплекса не приводила к росту концентрации данного цитокина выше показателей интактного контроля, что свидетельствовало об отсутствии у данного препарата провоспалительного действия.
Исследование изменения концентрации итерлейкина-1? (IL-1?) показало, что при внутрибрюшинной иммунизации чистым антигеном происходило умеренное дозозависимое повышение его концентрации.а Иммунизацияа термоденатурированной мономерной формой порина в составе ТИ-комплекса приводила к намного большему повышению уровня IL-1?. ТИ-комплекс в чистом виде вызывал умеренное повышение уровня IL-1?, демонстрируя способность обеспечивать более высокую функциональную способностьа иммунной системы к ответу на антигенное раздражение. Умеренное повышение уровня IL-1? под действием ТИ-комплексов свидетельствовало об отсутствии у ТИ-комплекса провоспалительных свойств.

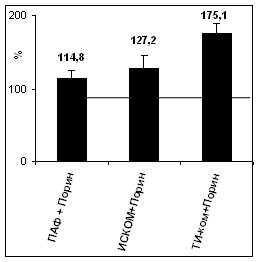
Рис. 7. Концентрация антител к термоденатурированной мономерной форме порина в сыворотке крови беспородных белых мышей при двукратной иммунизации индивидуальным порином и порином в составе ТИ-комплекса в дозах 0.1, 1 и 10 мкг/мышь. По оси ординат приведены показатели содержания антипориновых антител, определенных методом иммуноферментного анализа (ИФА) и выраженныха в единицах оптической плотности при длине волны 490 нм (разведение сыворотки 1:160). По оси абсцисс приведены экспериментальные группы животных.
аРис. 8. Различия в концентрации антител к термоденатурированной мономерной форме порина в сыворотке крови беспородных белых мышей после иммунизации данным антигеном в составе различных адъювантных носителей. По оси ординат приведены относительные концентрации антипориновых антител, выраженные в процентах к показателям сыворотки крови мышей, иммунизированных чистым белком (принят за 100%, показан горизонтальной линией). По оси абсцисс приведены экспериментальные группы животных.
Полученные данные об изменении концентрации цитокинов в сыворотки крови иммунизированных животных показали,а что ТИ-комплекс сам по себе обладал иммуномодулирующей активностью, являлся индуктором INF? и IL-1?.а
Эксперименты по оценке выживаемости мышей, инфицированных Y. pseudotuberculosis после иммунизации термоденатурированной мономерной формойа порина и порином в составе ТИ-комплексов, показали (рис.10), что иммунизация чистым антигеном в дозе 0.1 мкг на мышь приводила к увеличению средней длительности жизни в эксперименте вдвое, тогда как увеличение дозы приводило к снижению выживаемости животных. Иммунизация порином в составе ТИ-комплексов давала обратную картину: повышение показателей выживаемости животных при увеличении дозы. Таким образом, применение для иммунизации ТИ-комплексов в качестве носителя термоденатурированной мономерной формы порина приводила к устранению негативных свойств индивидуального антигена иа улучшению результатов вакцинации.
Полученные данные свидетельствовали о том, что ТИ-комплекс может служить эффективным носителем и адъювантом для термоденатурированной мономерной формы порина, не уступающим по свойствам наиболее сильным из известных адъювантов. Применение ТИ-комплекса при иммунизации животных порином позволяло не только повысить иммуногенность, но и ликвидировать иммуносупрессивные свойства данного антигена.
аа  аа
аа
Рис. 9. Концентрация INF? в сыворотке крови белых беспородных мышей иммунизированных внутрибрюшинно в дозах 0.1 и 1 мкг термоденатурированной мономерной формой порина и порином в составе ТИ-комплекса. По оси абсцисс приведены экспериментальные группы животных.
Рис. 10. Процент выживания и средняя длительность выживания мышей линии СВА, зараженных внутрибрюшинно 10 ЛД50 Yersiniapseudotuberculosis после иммунизации индивидуальным порином и порином в составе ТИ-комплексов в дозах 0.1, 1 и 10 мкг/мышь. По оси ординат приведен процент выживших животных по истечении 21 после заражения и асредняя продолжительность жизни мышей в эксперименте. По оси абсцисс - экспериментальные группы животных.
Оптимизацияа тубулярного иммуностимулирующего комплекса
Несмотря на наличие адъювантной активности, ТИ-комплекс КД-ХОЛ-МГДГ 3:2:6 обладал рядом недостатков: недостаточно высокой адъювантной активностью; результаты биоиспытаний оказывались невоспроизводимыми и часто противоречивыми; препарат носителя представлял собой весьма неоднородную суспензию; препарат тритерпеновых гликозидов, использованный для получения комплекса, был недостаточно охарактеризован химически, что также не могло способствовать получению воспроизводимых результатов.а
Оптимизация условий для формирования наночастиц тубулярного иммуностимулирующего комплекса
Для улучшения адъювантных свойств ТИ-комплекса мы решили заменить в его составе кукумариозид, на чистый кукумариозид А2-2. Использование чистого кукумариозида А2-2 в составе ТИ-комплекса предпочтительней использования суммы моносульфатированных тритерпеновых гликозидов, так как из всех тритерпеновых гликозидов из C. japonica именно кукумариозид А2-2 обладаета максимальной иммуностимулирующей активностью.
А
|
В |
Д
|
Б
|
Г |
Е |




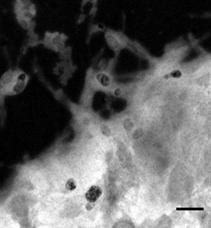
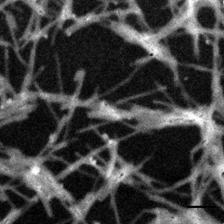
Рис. 11. Электронные микрофотографии структур, образованных системой КДА-ХОЛ-МГДГ с весовым соотношением компонентов А, Б - 3:2:6; В, Г - 1.5:2:6; Д, Е 6:2:4; А, В, Д - негативное контрастирование; Б, Г, Е - неокрашенный образец, EFTEM. Масштабная линия соответствует 100 нм.
Исследование препарата ТИ-комплекса КДА-ХОЛ-МГДГ 3:2:6 с использованием негативного контрастирования показало наличие типичных тубулярных частиц (рис. 11А). Однако, исследование препарата с помощью метода EFTEM (рис. 11Б) показало присутствие вещества, не входившего в состав тубулярных частиц, и представлявшего, вероятно, избыток не включившихся МГДГ и холестерола. Чтобы проверить это предположение, доля КДА в липид-сапониновой системе была сниженаа вдвое. В системе КДА-ХОЛ-МГДГ 1.5:2:6 резко возросло количество вещества, не вошедшего в состав тубулярных частиц, что хорошо видно на неокрашенных образцах (рис. 11Г). Тогда как на негативно контрастированных образцах обнаруживалось большое количество типичных тубул (рис. 11В).а Сравнение электронных микрофотографий систем КДА-ХОЛ-МГДГ 1.5:2:6 и 3:2:6 показывало, что вещество не входящее в состав тубулярных наночастиц представляло собой МГДГ и холестерол. В свою очередь, повышение доли кукумариозида А2-2 в липид-сапониновой системе вдвое,а КДА-ХОЛ-МГДГ 6:2:3 - приводило к формированию более однородной системы. На неокрашенных образцах обнаруживались небольшие количества вещества, не входившего в состав тубул. На негативно контрастированных образцах также были видны типичные тубулярные частиц. Дальнейшее снижение доли липидов в липид-сапониновой системе КДА-ХОЛ-МГДГ 6:2:4 приводило к формированию однородной системы, содержавшей только тубулярные частицы и практически не содержавшей постороннего вещества (видно на не окрашенных образцах, рис. 11Е). На негативно контрастированных образцах видны тубулярные частицы типичного строения (рис. 11Д). Повышение доли МГДГ в липид-сапониновой системеа КДА-ХОЛ-МГДГа 6:2:10 приводило к возрастанию количества вещества, не вошедшего в состав тубул в сравнении с системойа КДА-ХОЛ-МГДГа 6:2:6.
Анализ полученных микрофотографий показал, что для формирования тубулярных частиц в липид-сапониновых системах КДА-ХОЛ-МГДГ оптимальным являлось весовое соотношение компонентов 6:2:4. При этом важно, чтобы весовое соотношение КДА-ХОЛ было равным 6:2, тогда как весовое содержание МГДГ не обязательно должно быть постоянным и равным 4. Вполне допустимы ТИ-комплексы, в которых весовое соотношение КДА-ХОЛ-МГДГ равноа 6:2:4-6
Определение гемолитической активности оптимизированного тубулярного иммуностимулирующего комплекса
Замена кукумариозида на кукумариозид А2-2 в составе ТИ-комплекса, а также изменение соотношения компонентов, проведенное в результате оптимизации условий для его формирования, потребовало провести дополнительное исследование гемолитической активности ТИ-комплекса (табл. 2).а Ранее отмечалось, что из всех тритерпеновых гликозидов C.japonica кукумариозид А2-2 является самым активным гемолитиком, полученные данные подтверждали этоа (
Таблица 2.
Гемолитическая активность (EС50, мкг/мл) ТИ-комплексаа и его компонентов
|
EС50, мкг/мл |
Коэффициент ингибирования |
КДА |
0.78 |
- |
QuilA |
25 |
- |
МГДГZM |
44.24 |
- |
МГДГSP |
47.6 |
- |
6-2-15ZM |
13.6 |
17 |
6-2-15SP |
47.6 |
61 |
3-2-6ZM |
43 |
55 |
3-2-6SP |
60.84 |
78 |
6-2-4ZM |
71.9 |
92 |
6-2-4SP |
109.2 |
140 |
EС50 - концентрация вещества, вызывающая гемолиз 50% эритроцитов 0.5%-ной суспензии, , ZM - препараты, содержащие МГДГ из Zosteramarina, SP- содержащие МГДГ из Sargassumpallidum,соотношениекомпонентов ТИ-комплекса указано в порядке: КДА-ХОЛ-МГДГ.
Несколько неожиданным было обнаружение собственной гемолитической активности МГДГ, лишь вдвое меньшей, чем у сапонинов QuilA. Сравнение гемолитической активности ТИ-комплексов с различным составом показывало, что их гемолитическая активность повышалась при повышении доли МГДГ в комплексе и не росла при увеличении доли КДА. Сравнение гемолитической активности ТИ-комплекса КД-ХОЛ-МГДГ 3:2:6 и комплексом КДА-ХОЛ-МГДГ 6:2:4, полученным в результате оптимизации условий для формирования тубулярных частиц, показало, что оптимизированный комплекс также обладал меньшей гемолитической активностью. Примечательно, что оптимизированный ТИ-комплекс имел в 140 раз меньшую гемолитическую активность, чем исходный КДА.
Таким образом, на основании данных трансмиссионной электронной микроскопии и исследования гемолитической активности можно утверждать, что соотношение компонентов ТИ-комплексаа КД-ХОЛ-МГДГ 6:2:4 являлось оптимальным.
аУстойчивостьа тубулярного иммуностимулирующего комплекса
Для проведения биоиспытаний ТИ-комплекс КДА-ХОЛ-МГДГ 6:2:4 вводился мышам из расчета 1 мкг КД на 1 мышь в 200 мкл фосфатно-солевого буфера, что соответствовало 100 кратному разбавлению исходного препарата. Получаемые, в результате иммунизаций данные, имели широкий разброс и слабую воспроизводимость.
Исследование устойчивости ТИ-комплекса КДА-ХОЛ-МГДГ 6:2:4а показало, что при разбавлении фосфатно-солевым буфером происходило постепенное разрушение тубулярных наночастиц и увеличивалось количество вещества,а не входящего в состав тубул. При разбавлении в 5 раз признаков разрушения тубулярных частиц не наблюдалось; тогда как при разбавлении в 75 раз наблюдалось практически полное разрушение тубулярных наночастиц. Поэтому, с целью предотвращения разрушения ТИ-комплекса при введении его животным, следует как можно меньше разбавлять исходную липид-сапониновую суспензию, а потому необходимо иммунизировать животных минимально возможным объемом.а
Таким образом, в результате оптимизации состава ТИ-комплекса было установлено, чтоа оптимальным соотношением его компонентов, обеспечивающим минимальную токсичность и максимально эффективное формирование тубулярных наночастиц, исключающее наличие в препаратах, не вошедших в структуру частиц липидов, является КД-ХОЛ-МГДГ 6:2:4. Оптимальным способом введения животным оптимизированного ТИ-комплекса является разбавление исходного препарата с концентрацией липидов кукумариозидаа 1 мг/мл в 10 раз фосфатно-солевым буфером. Дальнейшее снижение разбавления комплекса невозможно, так как технически сложно иммунизировать животных объемами меньше 10 мкл.аа
Адъювантнаяа активность оптимизированного тубулярного иммуностимулирующего комплекса
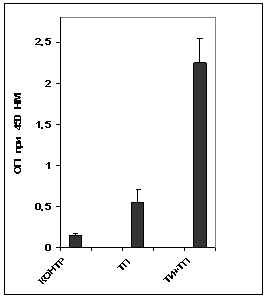
Анализ содержания специфических антител в сыворотке крови иммунизированных животных (рис. 12А) показал, что тример порина из наружной мембраны возбудителя псевдотуберкулеза Y. pseudotuberculosis в чистом виде умеренно иммуногенен. ТИ-комплекса КДА:ХОЛ:МГДГ 6:2:4 обеспечивал значительный адъювантный эффект, что выражалось в четырехкратном увеличении количества специфических антител в сыворотке крови животных иммунизированных порином в составе комплекса, в сравнении с чистым порином.
Анализа способности ТИ-комплекса изменять профиль цитокинов, вырабатываемыха в ходе иммунного ответа (рис.12 Б) показал, что иммунизация тримерным порином в составе ТИ-комплекса не приводила к воспалительной стимуляции (концентрация IL-12 росла незначительно, равно как и концентрация INF? не повышалась в сравнении с контролем). Вместе с тем, концентрация противовоспалительного цитокина IL-10 при иммунизации чистым порином незначительно падала, тогда как иммунизация порином с составе ТИ-комплекса приводила к меньшему падению его концентрации, что может свидетельствовать о способности оптимизированного комплекса снижать провоспалительную активность порина.
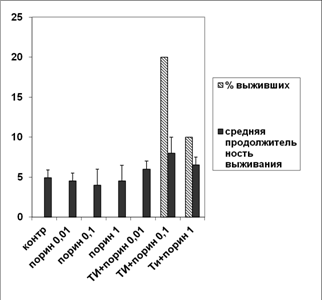
Определение протективного эффекта показало (рис. 13), что порин в чистом виде не защищал животных от гибели при экспериментальном псевдотуберкулезе, вне зависимости от дозы антигена.а Слабый протективный эффект наблюдался в группах животных, получивших порин в составе оптимизированного ТИ-комплекса.а При дозе порина 0.1 мкг на мышь наблюдался максимальный протективный эффект.
 а
а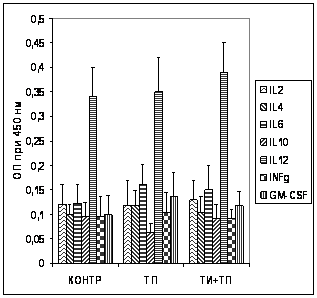
Б
Рис. 12. Содержание антипориновых антител (А) аи цитокинова (Б) в сыворотке крови мышей при двукратной иммунизации тримером поринаа и порином в составе ТИ-комплекса. По оси ординат - содержание специфических антипориновых антител и цитокинов, определенных методом иммуноферментного анализа и выраженных в единицах оптической плотности при длине волны 450 нм (разведение сыворотки крови 1/100). По оси абсцисс - экспериментальные группы животных. Обозначения: контр - интактный контроль, ТП - тример порина ТИ+ТП - тример порина в составе ТИ-комплекса.
аа
Рис. 13.а Выживаемость мышей линии СВА, зараженных 10LD50 Y. pseudotuberculosis. По оси ординат - средняя продолжительность выживания в эксперименте и процент выживших животных. По оси абсцисс - экспериментальные группы животных. Обозначения: контр - интактный контроль; порин 0.01, порин 0.1, порин 1 - порин в дозе 0.01, 0.1 и 1 мкг на мышь соответственно. ТИ+порин 0.01, ТИ+порин 0.1, ТИ+порин 1 а- порин в дозе 0.01, 0.1 и 1 мг на мышь соответственно, введенный в составе ТИ-комплекса на основе МГДГ из U. fenestrata
ВЫВОДЫ
1. Установлено, что моногалактозилдиацилглицерол (МГДГ) в отличие от других гликоглицеролипидов Laminariajaponica эффективно замещает фосфолипидную составляющую в классических иммуностимулирующих комплексах (ИСКОМ), не изменяя их везикулярной суперструктуры.
2. Показано, что сумма моносульфатированных тритерпеновых гликозидова изCucucmariajaponica - кукумариозид и кукумариозид А2-2, а также кукумариозид G1 из Eupentactafraudatrix, фрондозид Аа из Cucumariafrondosa и голотоксин А1 изApostichopusjaponicus формируют надмолекулярныеа комплексы с холестеролом в водной среде.а
3. Выявлено, что кукумариозид и кукумариозид А2-2 с холестеролома эффективно формируют трубчатые частицы, которые имеют четкие очертания иа характеризуются длиной тубул 150-350 нм, внешним диаметром 12-14 нм и диаметром канала 4 нм.
4. Найдено, что присутствие МГДГ в комплексах кукумариозида и голотоксина А1 с холестеролом необходимо для формирования гомогенных тубулярных частиц и снижения их гемолитической активности.
5. На примере порина из Yersiniapseudotuberculosisа и гемагглютинина вакцины Influvac показано, что комплекс кукумариозид-холестерол-МГДГ 3:2:6 может служить носителем субъединичных антигенов и высокоэффективным адъювантом, более эффективным, чем ИСКОМ и полный адъювант Фрейнда. Применение комплекса не приводит к развитию воспалительных реакций, снимает токсическое действие порина на систему интерферона, защищает 25% животных при экспериментальной инфекцииY. pseudotuberculosis.аа
6. Проведена оптимизация состава и процедуры полученияа ТИ-комплекса, путем замены суммы моносульфатированных тритерпеновых гликозидов из C. japonica на хроматографически чистый кукумариозид А2-2, и подбора соотношения его компонентов, чтоа позволило получать гомогенный препарат ТИ-комплекса без дополнительных процедур очистки.
7. Установлено, что оптимальным для формирования ТИ-комплекса является весовое соотношение компонентов кукумариозид А2-2-холестерол-МГДГ 6:2:4, обеспечивающее комплексу стабильно высокую адъювантную активность и протективность. Иммунизация тримером порина из Y. pseudotuberculosisв составе комплекса приводит к увеличению концентрации специфических антител в 4.5 раза, увеличивает среднюю продолжительность выживания в эксперименте и защищает 20% животных при экспериментальной инфекцииот этого возбудителя.а
8. Оптимизированный ТИ-комплекс обладает низкой гемолитической 109.2 мкг/мл, что соответствует 140-кратному снижению относительно чистого кукумариозида А2-2.
9. Разбавление препаратов ТИ-комплексаа фосфатно-солевым буфером приводит к разрушению тубулярных частиц; степень разрушения частиц возрастает с увеличением разбавления. Поэтому необходимо использовать для иммунизации исходный препарат, разбавленный не более чем в 10 раз.
Список работ по теме диссертации:
Статьи, опубликованные в ведущих рецензируемых научных журналах
- и И.А., Попов А.М.,а Костецкий Э.Я., Санина Н.М., Цыбульский А.М., Шныров В.Л., Мазейка А.Н., Богуславский В.М. Особенности мембранотропного действия некоторых тритерпеновых гликозидов голотурий, обладающих иммуностимулирующей активностью // Биофизика. 2008. Т. 53. с. 462-469.
- и И.А.,а Попов А.М.,а Цыбульский А.М., Санина Н.М., Костецкий Э.Я., Портнягина О.Ю., Новикова О.Д., Мазейка А.Н. Иммуностимулирующие свойства липид-гликозидного носителя антигена на основе кукумариозида А2-2 и моногалактозилдиацилглицерола из морской водоросли // Прикладная биохимия и микробиология. 2008. Т. 44.а № 6. C. 694-700.
- Мазейка А.Н., Попов А.М., Калинин В.И., Авилов С.А., Костецкий Э.Я. Комплексообразование тритерпеновых гликозидов голотурий как основа липид-сапониновых носителей субъединичных антигенов // Биофизика. 2008. Т. 53. № 5. С. 826-835.
- Kostetsky E.Y., Sanina N.M., Mazeika A.N., Tsybulsky A.V., Vorobyeva N.S., Shnyrov V.L. Tubular immunostimulating complex based on cucumarioside A2-2 and monogalactosyldiacylglycerol from marine macrophytes //Journal of Nanobiotechnology. 2011. 9:35.
- Цыбульский А.В., Попов А.М., Портнягина О.Ю., Артюков А.А., Санина Н.М., Мазейка А.Н., Новикова О.Д., Кривошапко О.Н., Костецкий Э.Я. Адъювантный эффект тубулярных иммуностимулирующих комплексов, модифицированных эхинохромом а,а в отношении порового белка из Yersiniapseudotuberculosis // Медицинская иммунология. 2011. Т. 13. № 2-3. С. 139-144.
- Цыбульский А.В., Попов А.М., Артюков А.А., Костецкий Э.Я., Кривошапко О.Н., Мазейка А.Н., Козловская Э.П.а Сравнительное изучение лечебного действия лютеолина, розмариновой кислоты и эхинохрома А при экспериментальной кардиопатологии, индуцированной стрессом // Биомедицинская химия. 2011. Т. 57. № 3. С. 314-325.
- Цыбульский А.В., Попов А.М., Санина Н.М., Мазейка А.Н., Портнягина О.Ю., Новикова О.Д.,а Тимченко Н.Ф., Костецкий Э.Я. Иммуногенные и протективные свойстваа наноразмерных конструкций на основе ТИ-комплексов и порообразующего белка из Yersinia pseudotuberculosis // Журнал микробиологии, эпидемиологии и иммунобиологии. 2011. № 2. С. 43-47.
- Sanina N.M., Kostetsky E.Y., Shnyrov V.L., Tsybulsky A.V., Novikova O.D., Portniagina O.Y., Vorobieva N.S., Mazeika A.N., Bogdanov M.V. The influence of monogalactosyldiacylglycerols from different marine macrophytes on immunogenicity and conformation of protein antigen of tubular immunostimulating complex // Biochimie. 2012. Vol. 94. № 4. P. 1048-1056.
аа Патенты:
- Костецкий Э.Я., Попов А.М., Санина Н.М., Ли И.А., Цыбульский А.В.,а Шныров В.Л., Мазейка А.Н. Носитель антигенов. Патент РФ № 2322259. Заявка № 2006123270/13 от 21.06.2006; опубл.20.04.2008 Бюл. №11.
- Костецкий Э.Я.,а Санина Н.М. , Мазейка А.Н., Цыбульский А.В., Воробъева Н.С., Новикова О.Д., Портнягина О.Ю., Шныров В.Л.а Иммуностимулирующий комплекс и его применение.а Заявка на патент № 2010115549 от 19.04.2010. Положительное решение о выдаче патента от 28.10.2011.
аа Работы, опубликованные в материалах региональных,а всероссийских и международных научных конференций и симпозиумах:
- и И.А., Попов А.М., Санина Н.М., Цыбульский А.В., Костецкий Э.Я., Портнягина О.Ю., Новикова О.Д., Мазейка А.Н., Богуславский В.М. Иммуностимулирующие свойства липид-гликозидного комплекса - носителя субъединичных антигенов // Исследования в области физико-химической биологии и биотехнологии: тез. докл. II региональной науч. конф. Владивосток, 2006. С. 119.
- и И.А., Попов А.М., Костецкий Э.Я., Санина Н.М., Мазейка А.Н., Богуславский В.М. Особенности мембранотропного действия тритерпеновых гликозидов морских голотурий, обладающих иммуностимулирующей активностью // Исследования в области физико-химической биологии и биотехнологии: тез. докл. II региональной науч. конф. Владивосток, 2006. С. 120.
- Мазейка А.Н., Попов А.М., Авилов С.А., Калинин В.И., Санина Н.М., Костецкий Э.Я., Портнягина О.Ю., Новикова О.Д. Комплексообразование тритерпеновых гликозидов голотурий с холестерином, как основа липид-сапониновых носителей белковых антигенов // XI международная молодежная школа-конференция по актуальным проблемам химии и биологии: сборник трудов. Владивосток 2007. С. 58.
- Воробъева Н.С., Мазейка А.Н. Модификация липид-сапониновых наноносителей белковых антигенов // Актуальные проблемы экологии, морской биологии и биотехнологии: IX региональная конференция студентов, аспирантов вузов и научных организаций Дальнего Востока России. Владивосток, 2010. С. 7.
- Mazeika A.N., Kostetsky, E.Ya., Sanina N.M. Hemolytiс activity of monogalactosyldiacylglycerol from marine macrophytes // Abst. of 3rd Asian Symposium on Plant Lipids. Yokohama, Japan, 2009. P. 321.
- Мазейка А.Н., Костецкий Э.Я., Воробьева Н.С., Цыбульский А.В. Иммуностимулирующая активность липид-сапонинового наноносителя антигенов на основе кукумариозида А2-2 // XIII Всероссийскаяа молодежная школа-конференция по актуальным проблемам химии и биологии: сб. трудов. Владивосток 2010. С. 39.
- Мазейка А.Н., Ли И.А., Попов А.М., Санина Н.М, Костецкий Э.Я., Портнягина О.Ю. Липид-сапониновые комплексы на основе тритерпеновых гликозидов голотурий и гликоглицеролипидов морских макрофитов - новый тип носителей антигенов для разработки субъединичных вакцин // X Международная молодежная Школа-конференция по актуальным проблемам химии и биологии: сб. трудов. Владивосток, 2006. С. 45.
- Sanina N., Kostetsky E., Tsybulsky A., Novikova O., Portniagina O., Mazeika A., Vorobieva N., Bogdanov M., Shnyrov V. Tubular immunostimulating complex based on lipids from marine hydrobionts // Abstr. of 52nd International Conference on the Bioscience of Lipids. Варшава, Польша, 2011. P. 22S.
- Sanina N., Kostetsky E., Shnyrov V., Novikova O., Portniagina O., Tsybulsky A., Bogdanov M., Mazeika A., Vorobyeva N. Influence of monogalactosyldiacylglycerol fromа marine macrophytes on immunogenici and conformation of protein antigen incorporated inа tubular immunostimmulating complex // Abstr. of 4th Asian Symposium on Plant Lipids. Hong Kong, 2011. P.10.
- Sanina N.M., Kostetsky E.Y., Shnyrov V.L., Novikova O.D., Portniagina O.Y., Tsybulsky, A.V., Bogdanov M.V., Mazeika A.N., Vorobieva N.S. Regulation of immunogenicity of protein antigen incorporated in tubular immunostimulating complex // Abstr. of 8th LIPIDOMICSа MEETING MEMBRANES and BIOACTIVE LIPIDS. Лион, Франция, 2011. P.64.
Мазейка Андрей Нколаевич
Разработка иммуностимулирующих комплексов на основе гликолипидов и ТРИТЕРПЕНОВЫХ ГЛИКОЗИДОВ из морских гидробионтов
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Подписано в печать 30 марта 2012-04-02. Формат 60Х80/16.
Усл. печ. л. 1.0 Уч. Изд.л. 1.0. Тираж 100 экз. Заказ №14
Отпечатано в типографии RICOH. Владивосток, ул.Фокина, 31. а
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]