
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
ВЛИЯНИЕ ТЕХНОГЕННОГО ЗАГРЯЗНЕНИЯ КОМБИНАТА СЕВЕРОНИКЕЛЬ НА РОСТ И РАЗВИТИЕ ДРЕВЕСНЫХ РАСТЕНИЙ (НА ПРИМЕРЕ BETULA CZEREPANOVII ORLOVA)
Автореферат кандидатской диссертации
На правах рукописи
УКИНА
Юлия Михайловна
ВЛИЯНИЕ ТЕХНОГЕННОГО ЗАГРЯЗНЕНИЯ КОМБИНАТА СЕВЕРОНИКЕЛЬ
НА РОСТ И РАЗВИТИЕ ДРЕВЕСНЫХ РАСТЕНИЙ
(НА ПРИМЕРЕ BETULA CZEREPANOVII ORLOVA)
03.02.08Цэкология
Автореферат диссертации на соискание ученой степени кандидата
биологических наук
Петрозаводск Ч 2011
Работа выполнена на кафедре биологии и химии
ФГБО УВПО Мурманский государственный гуманитарный университет
Научный руководительаа доктор биологических наук
доцент
Василевская Наталья Владимировна
Официальные оппоненты доктор биологических наук
профессор
Марковская Евгения Федоровна
доктор биологических наук
доцент
Чернобровкина Надежда Петровна
Ведущая организацияа Полярно-альпийский ботанический сад-
институт им. Н. А. Аврорина КН - РАН
Защита диссертации состоится л14 марта 2012 г. в 14 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете по адресу: 185910, Республика Карелия, г. Петрозаводск, пр. Ленина, 33, эколого-биологический факультет. E-mail: , факс: (88142)711000
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета, с авторефератом Ч на сайте ПетрГУ по адресу: Ученый секретарь
диссертационного совета,
канд. биол. наук И. М. Дзюбук
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы.Кольский полуостров является одним из наиболее индустриально развитых регионов Северо-Запада России. Экологическую ситуацию в регионе определяет концентрация крупнейших предприятий горнодобывающей промышленности и цветной металлургии, производственная деятельность которых в течение нескольких десятилетий привела к негативным последствиям для природных экосистем. Основным источником техногенного загрязнения окружающей среды в центральной части полуострова является крупнейший в Европе комбинат цветной металлургии ОАО Комбинат Североникель (г. Мончегорск).
Для оценки антропогенного воздействия с 60-х годов ХХ века начали использовать методы фитоиндикации (Крючков, 1976; Голубева, 1999; Черненькова, 2002). Накоплен большой фактический материал мониторинга техногенной среды с использованием различных критериев, таких как интенсивность физиологических процессов, уровень роста и развития морфологических и анатомических структур растений. Вокруг медно-никелевых комбинатов Североникель и Печенганикель проводится регулярный мониторинг состояния наземных экосистем, растительных популяций и сообществ, их продукционных характеристик (Крючков, 1982; Лукина, Никонов, 1996, 1998; Мазная, 2003; Kozlov, Zvereva, 2001, 2004). В то же время работы по изучению техногенного воздействия на динамику роста, развития и анатомическую структуру растений немногочисленны. В связи с этим, изучение онтогенетических реакций Betulaczerepanovii в условиях загрязнения среды тяжелыми металлами, является актуальным, поскольку позволяет выявить ответные реакции организма на техногенный стресс.
Цель исследования: изучение динамики процессов роста и развития Betula czerepanovii в условиях техногенного загрязнения комбината Североникель.
Задачи исследования:
- Изучить влияние техногенного загрязнения на динамику роста побегов и листьев, анатомическую структуру листа B. czerepanovii.
- Изучить влияние тяжелых металлов на процессы вегетативного развития B. czerepanovii.
- Изучить влияние тяжелых металлов на репродуктивное развитие B. czerepanovii и жизнеспособность семян.
Научная новизна. Впервые проведены исследования динамики морфометрических параметров B. czerepanovii в окрестностях комбината Североникель. Выявлены онтогенетические реакции процессов роста на увеличение содержания тяжелых металлов в почве по градиенту загрязнения в начале вегетационного сезона. Впервые изучена анатомическая структура листа B. czerepanovii в условиях техногенного загрязнения медно-никелевых производств. Показано, что ионы никеля и меди вызывают более значительную модификацию палисадных клетокмезофилла B. czerepanovii, выражающуюся в их удлинении, уменьшении диаметра, увеличении индекса палисадности, усилении периклинальных делений. В меньшей степени изменяется величина губчатых клеток. Впервые исследованы динамика числа листьев на побеге, скорость их развертывания, органогенез почек B. czerepanovii. Установлено, что техногенное загрязнение ингибирует развитие листьев и скорость их развертывания на побеге текущего года в первые дни вегетации в окрестностях комбината, но не влияет на процессы дифференциации апикальных меристем и число листьев в почках. Получены новые данные по репродуктивному развитию березы, произрастающей в окрестностях комбината. Выявлено, что по градиенту загрязнения ионами никеля и меди отмечается усиление нестабильности репродуктивного развития B. czerepanovii, что выражается в появлении особей, продуцирующих аномально большое число только пестичных или тычиночных соцветий, а также особей, полностью утративших репродуктивную функцию. Эксперименты показали, что растения, выращенные из семян B. czerepanovii техногенной пустоши и редколесья нежизнеспособны, у них выявлены замедление развития, хлороз и пожелтение листьев, ранняя дефолиация, наличие большой доли тератов, что связано с низкой жизнеспособностью семян, сформированных в условиях высокой концентрации никеля и меди в техногенной среде.
Практическая значимость. Результаты исследований могут быть использованы для оценки состояния популяции B. czerepanovii в окрестностях комбината Североникель, разработки системы мероприятий по восстановлению техногенных пустошей и проведения биомониторинга лесных сообществ. Данные по стабильности развития популяции B. czerepanovii на основании индекса флуктуирующей асимметрии в окрестностях комбината Североникель могут быть использованы для оценки состояния природной среды и организации мониторинговых исследований на территории г. Мончегорска. Результаты исследований могут быть использованы в курсах лекций Экология, Биомониторинг окружающей среды для студентов биологических и экологических специальностей университетов.
Апробация работы. Основные положения работы были представлены на 10 научных конференциях различного уровня, включая пять международных конференций: Экологические проблемы северных регионов и пути их решения (Апатиты, 2004), Устойчивость экосистем и проблема сохранения биоразнообразия на Севере (Кировск, 2006), Экологическое образование и экологическая наука для устойчивого развития (Архангельск, 2007), Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт (Мурманск, 2009), Промышленная ботаника: состояние и перспективы развития (Донецк, 2010); две всероссийские конференции с международным участием: Проблемы региональной экологии в условиях устойчивого развития (Киров, 2007), Экологические проблемы промышленных городов (Саратов, 2011); две всероссийские конференции: Ботанические проблемы регионального природопользования (Рязань, 2001), Принципы и способы сохранения биоразнообразия (Пущино, 2008); научно-практическую конференцию студентов и аспирантов Естественнонаучные проблемы арктического региона (Мурманск, 2003).
Публикации. По теме диссертации опубликовано 15 работ, в том числе две статьи в журналах, рекомендованных ВАК РФ.
Организация, объем исследований и личный вклад автора. Работа выполнена на кафедре биологии и химии Мурманского государственного гуманитарного университета в 2006Ц2011 гг. Все полевые работы, обработка, обобщение и анализ материалов исследований проведены автором.
Структура и объем диссертации. Диссертация состоит из введения, пяти глав, заключения, выводов, списка литературы (344 источника, в том числе 80 на иностранных языках) и приложения. Работа изложена на 172 страницах, содержит 11 таблиц и 49 рисунков.
Благодарности. Автор выражает глубокую благодарность своему научному руководителю Ч д-ру биол. наук, профессору кафедры биологии и химии МГГУ Н. В. Василевской за разностороннюю помощь и постоянную поддержку, советы и конструктивные замечания при подготовке диссертационной работы. Искренне признательна всем сотрудникам кафедры биологии и химии МГГУ за доброжелательное отношение и участие. Выражаю признательность заслуженному учителю Российской Федерации К. Н. Сорбалэ, научному сотруднику Лапландского государственного биосферного заповедника Н. В. Зануздаевой. Благодарю зав. лаборатории наземных экосистем Института проблем промышленной экологии Севера, к.б.н. Л. Г. Исаеву за помощь при проведении исследований. Благодарю членов моей семьи за поддержку на всех этапах исследований.
СОДЕРЖАНИЕ РАБОТЫ
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
1.1 Воздействие техногенного загрязнения на процессы роста растений
В разделе приводится анализ публикаций, монографий, обзорных и оригинальных статей отечественных и зарубежных авторов, в которых рассматривается воздействие газообразных токсикантов и тяжелых металлов на растения в условиях техногенного стресса. Рассмотрены различные аспекты физиологической роли и токсического действия никеля и меди на растения.
1.2 Влияние поллютантов на рост листьев и формирование
фотосинтетического аппарата
В разделе приведены литературные данные по многообразию симптомов повреждений древесных видов растений. Рассмотрено влияние техногенного загрязнения среды на процессы роста и анатомо-физиологические параметры листьев. Описаны результаты анатомических исследований мезоструктуры листа, которые используются для диагностики повреждений растений промышленными выбросами.
1.3 Влияние техногенного загрязнения на процессы вегетативного развития растений
Рассматриваются данные по воздействию тяжелых металлов на апикальные меристемы высших растений, типы морфологических аномалий развития. Представлен обзор литературы отечественных и зарубежных авторов по флуктуирующей асимметрии, которая является одним из показателей нарушения развития растений в условиях техногенной среды.
1.4 Влияние техногенного загрязнения на процессы
репродуктивного развития растений
Приводится обзор работ, в которых выявлено воздействие техногенного загрязнения на цветение и плодоношение, качество и количество семян, жизнеспособность семенного потомства. Обсуждаются результаты изучения репродуктивного развития ряда видов деревьев и кустарничков в условиях техногенного загрязнения комбината Североникель.
ГЛАВА 2. ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
2.1 Объект исследований
Исследования роста и развития выполнены на березе Черепанова (Betulaczerepanovii Orlova) Ч виде, широко распространенном в условиях Европейской Субарктики. B. czerepanovii Ч гипоарктический, евросибирский вид. Вопросы распространения и таксономии рода Betula на Кольском полуострове являются дискуссионными. Важной особенностью представителей рода является их высокая морфологическая пластичность, создающая значительные сложности в таксономических исследованиях (Gardiner, 1984). В. Н. Сукачев (1921) предполагал, что все описываемые виды Betula Ч это климатические формы B. pubescens. В. И. Ермаков (1986) высказал предположение, что основные виды рода Betula Ч B. pubescens и B. pendula представлены на Кольском полуострове северными расами. Н. И. Орлова (1978) выделила вид Betula czerepanovii Orlova. По ее мнению, данный вид Ч гибрид между B. pubescens Ehrh. и B. nana L. Ряд западных ученых определяют исследуемый вид березы в окрестностях комбината Североникель как Betulapubescens subsp. czerepanovii Orlova Hamet-Ahti. (Neuvonen et al., 1991; Ruohomaki et al., 1996). Мы рассматриваем изучаемый вид как Betula czerepanovii Orlova. В зоне техногенного воздействия комбината Североникель B. czerepanovii присутствует во всех типах сообществ, вне зависимости от степени их нарушенности, выживает на техногенных пустошах, где приобретает низкорослую форму с множеством стволов (Kryuchkov, 1993).
2.2 Методы исследований
Исследования проводили в полевые сезоны 2005Ц2007 гг. на пробных площадях, расположенных по градиенту загрязнения в юго-восточном направлении от комбината Североникель (г. Мончегорск, Мурманская область). Экспериментальные площади заложены в техногенной пустоши на расстоянии 3 км от источника загрязнения (ПП1), зонах техногенного редколесья (8 км Ч ПП2) и дефолиирующих лесов (20 км Ч ПП3 и 30 км Ч ПП4). Фоновая площадка (контроль Ч ПП5) располагалась в 60 км от комбината. На всех модельных площадках выполнены геоботанические описания растительности (МетодыЕ, 2002). Изучение содержания тяжелых металлов в почве и листьях B. czerepanovii проводили методом атомно-абсорбционной спектрофотометрии в лаборатории наземных экосистем Института проблем промышленной экологии Севера КН - РАН (г. Апатиты).
На каждой пробной площади маркировали 10 близких по габитусу деревьевB. сzerepanovii, в средней части кроны отмечали по 5 побегов. В течение полевых сезонов 2005Ц2006 гг. измеряли длину годичного прироста, число развернувшихся листьев, длину и ширину нижних листьев. Исследования проводили в динамике: с конца мая до начала августа. На основании данных по динамике числа листьев на побеге, рассчитана скорость развертывания листьев, под которой мы понимаем среднее количество суток, необходимое для развертывания одного листа на побеге текущего года. При этом число суток (от предыдущего учета числа листьев на приросте до следующего учета) делили на число развернувшихся листьев за данное время. В начале августа 2006 г. собраны листья березы для изучения их анатомической структуры. В качестве фиксатора использован FAA. На поперечных срезах измерялись показатели: длина и ширина палисадной и губчатой клетки, число слоев, толщина палисадного и губчатого мезофилла, толщина листовой пластинки. Индекс палисадности рассчитывали как отношение толщины палисадного мезофилла к толщине губчатого (Горышина, 1989). Для изучения влияния техногенного загрязнения на дифференциацию апикальных меристем и заложение листьев, в мае 2005Ц2006 гг. на всех площадках собраны терминальные почки с ауксибластов B. czerepanovii, которые фиксировали в 50% спирте. Для исследования структуры почек применен морфо- физиологический анализ по Ф. М. Куперман (1982).
При изучении стабильности развития популяции B. czerepanovii использован метод флуктуирующей асимметрии. Сбор и гербаризация листьев проводились в конце августа, общая биологическая повторность составила 100 листьев с каждой площадки. Величина флуктуирующей асимметрии оценивалась по формуле: FA = 2 ? |WL ? WR| / (WL + WR) (по Palmer, Strobeck, 1986), где WL Ч ширина левой половины листа, WR Ч ширина правой половины листа. В числителе разность берется по модулю.
Для изучения показателей репродуктивного развития B. czerepanovii в течение трех полевых сезонов (2005Ц2007 гг.) в середине июня проводили учет мужских и женских сережек. Биологическая повторность репродуктивных побегов на каждой площадке 50-кратная. С этих же деревьев в конце августа 2005 г. собраны семена для проведения лабораторных экспериментов. Семена стратифицировали в снегу в течение 45 суток, затем высаживали в сосуды с дерново-подзолистой почвой. Проращивание семян проводилось в лабораторных условиях, в теплице при постоянной температуре воздуха 20Ц23С, влажности 60Ц70%, спектральный состав света соответствовал облучению лампами ЛБУ-30. Всего заложено 5 вариантов опыта: 1 вариант Ч техногенная пустошь, 2 Ч техногенное редколесье, 3 Ч дефолиирующий лес (20 км), 4 Ч дефолиирующий лес (30 км), 5 Ч контроль (фоновая зона). Появившиеся всходы выращивали в течение 80 суток, при этом в течение 45 суток (майЦиюнь 2006 г.) проводили исследования динамики роста и развития растений. Учет проростков проводили на 3, 5, 7, 10, 15 день эксперимента (Семена древесныхЕ, 1972). Определяли грунтовую всхожесть и энергию прорастания. Линейные размеры проростков и ювенильных растений измеряли на 5, 7, 15, 45-й день после высева семян. Отмечали число погибших и поврежденных особей, нарушения развития.
Статистическая обработка полученных результатов выполнена на основе дисперсионного и корреляционного анализа с помощью стандартного пакета программ Microsoft Excel 2003. Взаимосвязь показателей роста и развития с содержанием никеля и меди в почве и листьях оценивали при помощи коэффициентов корреляции. Достоверность различий между средними арифметическими оценивалась по t-критерию Стьюдента, различия считались статистически значимыми при p < 0,05.
ГЛАВА 3. ПРИРОДНО-КЛИМАТИЧЕСКИЕ УСЛОВИЯ
РАЙОНА ИССЛЕДОВАНИЯ
Глава состоит из семи разделов, в которых рассматриваются физико-географическое положение и рельеф, геология, климат, почвы и растительность, техногенное загрязнение среды.
Исследования проводили в центральной части Кольского полуострова, на территории, относящейся к подзоне северо-таежных лесов Карельско-Кольской подпровинции Североевропейской таежной провинции (ГеоботаническоеЕ, 1989) в зоне загрязнения комбината Североникель (г. Мончегорск). Город Мончегорск расположен на 68 северной широты и 32,5 восточной долготы. Комбинат Североникель и г. Мончегорск с востока ограничены озером Имандра и Хибинским горным массивом, с запада Ч горной системой Волчьих, Монче и Чуна-тундр. В окрестностях г. Мончегорска средняя температура зимой составляет от ?9С до ?14С, средняя температура в период с мая по сентябрь от 7,5С до 10,6С. Сумма температур вегетационного периода Ч 1300С. Количество дней с температурой более 5С составляет 109Ц130.
Мончегорский район является наиболее загрязненной территорией Мурманской области. Зона очень опасного уровня загрязнения составляет площадь 1400 км2 и характеризуется максимальными уровнями годовых выпадений Ni и Cu (115,9 и 136,5 кг/км2, соответственно) (Мокротоварова, 2003).
Одним из основных источников промышленного загрязнения является медно-никелевый комбинат Североникель Ч один из крупнейших производителей никеля в мире. В настоящее время в качестве сырья комбинат использует флотационный медно-никелевый концентрат (Ni Ч 4%, Cu Ч 2,5%) Норильского месторождения. На разных этапах технологического цикла в атмосферу поступают металлы в виде сульфидов, не полностью окисленных сульфидов, окисей и закисей, а также пыль, содержащая тяжелые цветные металлы: никель, медь, кобальт. Значительная часть металлов переносится воздушным путем в твердом дисперсном состоянии на большие расстояния. В связи с преобладанием северных и южных ветров загрязненная территория имеет форму эллипса. Площадь зоны загрязнения в конце XX века составляла до 93 000 га, причем на площади 3,7 тыс. га хвойные леса уничтожены полностью (Poznyakov, 1993).
ГЛАВА 4. ВЛИЯНИЕ ТЕХНОГЕННОГО ЗАГРЯЗНЕНИЯ НА ДИНАМИКУ РОСТА BETULA CZEREPANOVII В ОКРЕСТНОСТЯХ КОМБИНАТА СЕВЕРОНИКЕЛЬ
4.1 Содержание тяжелых металлов и серы в почве
В зоне техногенного воздействия комбината Североникель выявлено увеличение содержания тяжелых металлов (Ni, Cu) в почве по градиенту загрязнения, эта зависимость имеет нелинейный характер. В 2006 г. на расстоянии 60 км от комбината (ПП5) отмечается превышение ПДК по никелю в 3 раза, в образцах почвы дефолиирующих лесов (ПП4, ПП3) в 5Ц6 раз, в зоне техногенного редколесья (ПП2) Ч в 19 раз выше нормы. Резкое увеличение содержания никеля в почве Ч в 198 раз зарегистрировано на техногенной пустоши (ПП1). Накопление меди в почве имеет иной характер. В фоновой зоне ее концентрация превышает норму в 6 раз, на ПП4, ПП3 в 13 и 19 раз (соответственно), а на ПП2 содержание меди достигает максимума и в 74 раза превышает ПДК. Содержание меди в образцах почвы на техногенной пустоши превышает ПДК в 50 раз.
4.2 Воздействие техногенного загрязнения на состояние растительности
в окрестностях комбината Североникель
Выявлены существенные изменения состава и уменьшение числа видов лихено-бриологической группы на площадках техногенной пустоши и редколесья. При этом есть виды сосудистых растений, адаптированные к условиям хронического загрязнения, которые выдерживают высокие концентрации тяжелых металлов и газообразных поллютантов и способствуют формированию устойчивых сообществ. Такими видами являются: Betula czerepanovii, Salix borealis, Empetrum hermaphroditum и представители рода Vaccinium, которые произрастают на всех пробных площадях в условиях техногенной нагрузки вне зависимости от степени нарушенности экотопов.
4.3 Динамика роста побегов Betula czerepanovii в условиях техногенного загрязнения
Мониторинг динамики роста побегов текущего года B. czerepanovii показал, что прирост побегов значительно уменьшается по градиенту загрязнения, этот эффект более выражен в начале вегетации. Особенно значительное влияние техногенного загрязнения на рост побегов выявлено в конце мая в зоне техногенного редколесья (ПП2) и техногенной пустоши (ПП1): в 2,7 и 3,5 раза (соответственно) меньше фоновых значений в 2005 г., в 3,2 и 3,5 раза в 2006 г. (рис. 1). Через 5Ц10 дней ингибирование ослабевает, длина приростов меньше контрольных значений в 1,9 и 2,6 раза (соответственно). Зависимость длины побега от содержания никеля и меди в почве пробных площадей в 2006 г. подтверждается данными корреляционного анализа (для никеля R = ?0,90Ц0,97, p < 0,05; для меди R = ?0,89Ц0,96, p < 0,05). Анализ показал, что при повышении содержания никеля и меди в почве, длина побега текущего года уменьшается.
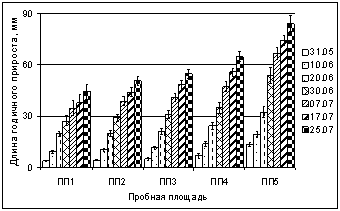
Рис. 1. Влияние техногенного загрязнения на динамику приростов
Betulа czerepanovii в окрестностях комбината Североникель в 2006 г. (в мм)
Известно, что ионы тяжелых металлов оказывают негативное воздействие на клеточное деление (Серегин, Кожевникова, 2006). Наблюдаемое ингибирование роста побегов B. czerepanovii в условиях техногенной нагрузки в начале вегетации свидетельствует, по-видимому, о том, что ионы никеля и меди оказывают более значительное токсическое воздействие на процессы митотических делений клеток в апексах побеговв начале их ростаи меньшее на последующее деление и растяжение клеток. Ряд авторов также указывают на тот факт, что техногенная среда оказывает влияние на постэмбриональную стадию побегообразования у древесных растений (Полевой и др., 1991; Васфилов, 2003).
4.4 Динамика роста листьев Betula czerepanovii в условиях техногенного загрязнения
4.4.1 Содержание тяжелых металлов в листьях Betula czerepanovii
Исследования показали, что концентрации Ni и Cu в листьях B. czerepanovii многократно увеличиваются по градиенту загрязнения (табл. 1). Для сопоставления полученных данных в качестве фоновой выбрана территория, которая соответствует критериям, принятым в международной практике (Кислотные осадки.., 1999) и расположенная на значительном (250 км) удалении от комбината Североникель в юго-западной части Мурманской области (Отчет о НИР ИППЭС Е, 2003).
Таблица 1
Содержание меди и никеля в листьях Betula czerepanovii
в окрестностях комбината Североникель в 2005Ц2006 гг. (мг/кг)
№ пробной площади |
Ni мг/кг |
Cu мг/кг |
||
2005 г. |
2006 г. |
2005 г. |
2006 г. |
|
ПП1 |
171,8 |
191,6 |
26,5 |
39,4 |
ПП2 |
227,2 |
169,3 |
44,9 |
50,2 |
ПП3 |
101,9 |
79,6 |
30,5 |
28,6 |
ПП4 |
37,6 |
45,6 |
13,4 |
19 |
ПП5 |
29,3 |
26,9 |
14,8 |
12,9 |
фон* |
4,5 |
Ч |
5 |
Ч |
*Приведены значения концентрации никеля и меди в листьях березы для фоновой площади в пос. Алакуртти (250 км от г. Мончегорска) (Отчет о НИР ИППЭС КН - РАНЕ, 2003).
4.4.2 Динамика роста листьев
В результате мониторинга динамики роста листьев березы получено, что в условиях высокой техногенной нагрузки на площадках ПП3, ПП2, ПП1 происходит ингибирование роста листьев, особенно в первые дни вегетации (рис. 2). В начале развертывания в конце мая 2005 г. длина листа в зоне техногенного редколесья (ПП2) в 1,3 раза, в 2006 г. в 1,7 раза меньше фоновых значений (ПП5). На техногенной пустоши (ПП1) этот показатель Ч в 1,5 (2005 г.) и 1,7 раза (2006 г.) меньше, чем в фоне. Через 10 дней после начала роста листьев эти различия снижается на ПП1 и ПП2 до 1,2Ц1,3 раза, что сохраняется в течение всего периода наблюдений.
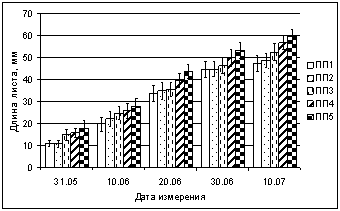
Рис. 2. Влияние техногенного загрязнения на длину листа Betula czerepanovii
в окрестностях комбината Североникель в 2006 г. (мм)
При изучении техногенного воздействия на динамику роста листьев березы в ширину также выявлено уменьшение этого показателя по градиенту загрязнения (рис. 3). В зоне техногенного редколесья (ПП2) в начале развертывания ширина листа в конце мая 2005, 2006 гг. в 1,4 и 1,7 раза (соответственно) меньше фоновых значений, на техногенной пустоши (ПП1) этот показатель Ч в 1,6 (2005 г) и 1,8 раза (2006 г.) меньше фона. Через 10 дней после начала роста листьев эти различия снижаются. Корреляционный анализ данных за 2005 и 2006 гг. выявил достоверную зависимость между содержанием в листьях никеля и меди и размерами листа. Получены высокие статистически значимые (p < 0,05) отрицательные корреляции. В 2006 г. коэффициенты зависимости длины листа (в динамике) от содержания никеля R = ?0,87Ц0,98, меди R = ?0,86Ц0,95. Коэффициенты корреляции ширины листа от содержания никеля R = ?0,90Ц0,98, меди R = ?0,81Ц0,96. Результаты анализа показали, что при повышении содержания никеля и меди в листьях, их длина и ширина достоверно уменьшается.
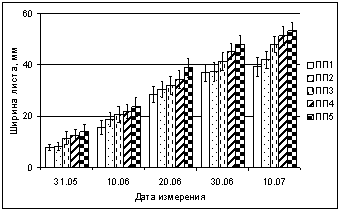
Рис. 3. Влияние техногенного загрязнения на ширину листа Betula czerepanovii
в окрестностях комбината Североникель в 2006 г. (мм)
Таким образом, низкие показатели длины и ширины листьев B. czerepanovii в начале вегетации обусловлены высокими концентрациями тяжелых металлов. По-видимому, ионы никеля и меди оказывают большее ингибирующее влияние на процессы деления клеток апикальных меристем в начальный период внепочечного развития побега и листьев, когда процессы дифференцировки клеток наиболее интенсивны. В результате замедления процессов развития, происходит торможение роста. Результаты наших исследований согласуются с данными ряда авторов (Таланова и др., 2001; Sandalio et al., 2001; Khudsar et al., 2004; Kosobrukhov et al., 2004; Лайдинен и др., 2004), которыми также получено, что повышение концентрации тяжелых металлов в окружающей среде приводит к значительному уменьшению длины, ширины и площади листовой пластинки, что является одной из причин снижения интенсивности процесса фотосинтеза.
4.5 Влияние техногенного загрязнения на анатомическую
структуру листа Betula czerepanovii
Исследования количественных показателей мезоструктуры листа выявили, что параметры клеток изменяются по градиенту загрязнения. Палисадные клетки значительно удлиняются: на 65% (ПП2), на 100% (ПП1) по сравнению с фоном и уменьшаются в диаметре по градиенту загрязнения (табл. 2). За счет роста клеток в длину увеличивается толщина палисадного мезофилла: на 20% (ПП3), 30% (ПП2), 66% (ПП1). Длина губчатых клеток листьев березы увеличивается на 47% (ПП3), 40% (ПП2), 53% (ПП1), соответственно возрастает толщина губчатого мезофилла. Достоверно уменьшается диаметр клеток губчатой ткани на площадках ПП1 и ПП2. По мере возрастания техногенной нагрузки увеличивается толщина листовой пластинки:на 50% (ПП2, ПП1) по сравнению с контролем. Корреляционный анализ выявил статистически значимые (p < 0,05)сильныекорреляции между содержанием в листьях тяжёлых металлов и анатомическими параметрами. По данным 2006 г. получены высокие положительные коэффициенты корреляции между длиной палисадной клетки и содержанием в листьях никеля (R = 0,91) и меди (R = 0,72), а также между длиной губчатой клетки и концентрацией тяжелых металлов (никель R = 0,76; медь R = 0,71). Между диаметром (шириной) палисадных и губчатых клеток и содержанием никеля и меди в листьях выявлены высокие отрицательные значения коэффициентов корреляции. Чем больше содержание никеля и меди в листьях, тем меньше диаметр палисадных клеток (R = ?0,93; R = ?0,81, соответственно). Аналогичная зависимость получена по ширине губчатых клеток (никель R = ?0,92; медь R = ?0,72). Толщина листовой пластинки и содержание никеля (R = 0,93) и меди (R = 0,93) имеют высокую положительную корреляционную зависимость, то есть чем больше тяжелых металлов накапливается в листьях, тем интенсивнее лист растет в толщину.
Таблица 2
Влияние техногенного загрязнения на анатомическую структуру листьев
Betula czerepanovii в окрестностях комбината Североникель в 2006 г. (в мкм)
Показатели (N = 100) |
ПП1 |
ПП2 |
ПП3 |
ПП4 |
ПП5 |
Толщина листа |
25,91,7 |
25,61,5 |
23,51,4 |
18,41,1 |
16,91,7 |
Толщина палисадного мезофилла |
9,81,4 |
7,71,5 |
7,11,1 |
5,61,0 |
5,90,8 |
Число слоев палисадного мезофилла |
1,80,3 |
1,40,3 |
1,50,4 |
1,30,3 |
1,30,2 |
Толщина губчатого мезофилла |
11,21,6 |
11,01,6 |
10,31,1 |
8,91,0 |
8,71,6 |
Число слоев губчатого мезофилла |
5,00,8 |
5,60,8 |
6,00,6 |
5,20,7 |
5,60,8 |
Длина палисадной клетки |
4,60,9 |
3,80,7 |
2,90,6 |
2,20,5 |
2,30,5 |
Ширина палисадной клетки |
0,70,2 |
0,90,4 |
1,00,2 |
1,10,4 |
1,20,5 |
Длина губчатой клетки |
2,30,3 |
2,10,5 |
2,20,5 |
1,90,5 |
1,50,4 |
Ширина губчатой клетки |
0,90,3 |
1,20,3 |
1,30,4 |
1,40,3 |
1,40,4 |
Индекс палисадности |
0,9 |
0,7 |
0,7 |
0,6 |
0,7 |
ДПМ/ШПМ* |
6,6 |
4,2 |
3 |
1,9 |
1,9 |
*Соотношение длины клетки палисадного мезофилла к ширине клетки.
Известно, что тяжелые металлы, в частности, никель вызывают анатомические изменения. Согласно полученным нами данным высокая концентрация никеля и меди в листьях вызывает более значительную модификацию палисадных клетокмезофилла B. czerepanovii, выражающуюся в их удлинении, уменьшении диаметра, увеличении индекса палисадности, усилении периклинальных делений. Увеличение размеров клеток мезофилла и толщины листа, по-видимому, является неспецифической структурной реакцией B. czerepanovii на техногенное загрязнение среды. Аналогичные данные получены для рода Betula Ю. В. Гамалеем (2004) в экстремальных условиях произрастания, что, как считает автор, свидетельствует о неспецифической реакции ассимиляционного аппарата берез на действие стрессовых факторов.
ГЛАВА 5. ВЕГЕТАТИВНОЕ И РЕПРОДУКТИВНОЕ РАЗВИТИЕ
BETULA CZEREPANOVII В УСЛОВИЯХ ТЕХНОГЕННОГО ЗАГРЯЗНЕНИЯ КОМБИНАТА СЕВЕРОНИКЕЛЬ
5.1 Повреждение листьев Betula czerepanovii
Исследования показали, что с усилением техногенного загрязнения происходит увеличение числа поврежденных листьев. Начальное повреждение листьев отмечено на всех пробных площадях. Оно выражалось в появлении точечности, некрозе кончика и края листа, мумификации части листьев, преждевременном появлении осенней окраски. На техногенной пустоши и зоне техногенного редколесья в начале июля часть листьев B. czerepanovii приобретает коричневый цвет, здоровые листья практически отсутствуют. На этих пробных площадях резко увеличивается число листьев с максимальной степенью повреждения в виде хлорозных и некрозных пятен, усыхания отдельных листьев или их скручивании, по сравнению с фоновыми условиями. У особей B. czerepanovii, произрастающих в непосредственной близости к комбинату, происходит быстрое старение листьев, что выражается в снижении продолжительности их вегетации за счет преждевременной дефолиации (20.08.2005; 25.08.2006), тогда как начало листопада в лесах фоновой зоны по средним многолетним данным Лапландского заповедника приходится на вторую декаду сентября.
На трех пробных площадках (ПП3, ПП2, ПП1) выявлены массовые энтомоповреждения молодых листьев мелкими насекомыми Ч листоблошками или медяницами (Сем. Psyllidae), тлями (Сем. Aphididae). Аналогичные данные по значительному повреждению листьев деревьев вредителями получены Е. Л. Зверевой и М. В. Козловым для Salix borealis и S. сaprea в окрестностях комбината Североникель (Zvereva, Kozlov, 2001). В зоне техногенного редколесья и дефолиирующих лесов наблюдается раннее пожелтение листьев березы, которое вызывает ржавчинный гриб Melampsoridium betulinum Kleb.
5.2 Развитие листьев Betula czerepanovii в условиях техногенного загрязнения
5.2.1 Число листьев
В ходе исследований установлено, что в 2005 г. на пробных площадках, расположенных в фоновой зоне и дефолиирующих лесов развертывание листьев началось на 5 суток ранее (22.05Ц23.05), чем у особей B. czerepanovii, произрастающихна ПП2 и ПП1(28.05). В 2006 г. в фоновой зоне развертывание листьев зафиксировано 25.05, на техногенной пустоши 30.05. Таким образом, в условиях острого загрязнения среды ионами никеля и меди начало внепочечного развития листьев березы задерживается на 5 суток. При изучении динамики числа листьев B. czerepanovii на побегах текущего года вокрестностях комбината Североникель получено, что в зоне дефолиирующих лесов (ПП4, ПП3) число листьев B. czerepanovii в первые дни вегетации в 1,9 (30.05.2005) и 1,3 раза (31.05.2006) меньше, чем в фоновых условиях. Уже через пять дней этот эффект снижается до 1,2 раза и сохраняется до конца июля. Наиболее значительное ингибирование развития листьев выявлено на площадках ПП2 и ПП1. В начале вегетации (10.06.06Ц20.06.06) число листьев в зоне техногенного редколесья в 1,9Ц1,5, на техногенной пустоши в 2Ц1,7 раза меньше по сравнению с фоновыми значениями. Начиная с конца июня ингибирование развития листьев становится менее выраженным и снижается до 1,3Ц1,4 раза, что сохраняется до конца июля. Аналогичные данные получены и в 2005 г.
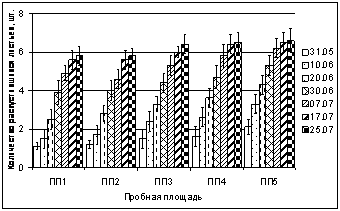
Рис. 4. Влияние техногенного загрязнения на динамику распускания листьев
Betula czerepanovii в окрестностях комбината Североникель в 2006 г. (шт.)
Одной из причин уменьшения числа листьев на побегах B. czerepanovii в окрестностях комбината Североникель является наличие в них тяжелых металлов в концентрациях значительно превышающих ПДК. По данным 2006 г. получены высокие отрицательные коэффициенты корреляции между числом листьев на побеге и содержанием в листьях никеля (R = ?0,92Ц0,99, р < 0,05) и меди (R = ?0,89Ц0,99, р < 0,05): при повышении содержания в листьях никеля и меди, число листьев на побеге текущего года уменьшается. Таким образом, данные по динамике листьев подтверждают, что высокие концентрации ионов никеля и меди ингибируют дифференциацию клеток в апикальных меристемах листьев B. czerepanovii по градиенту загрязнения, особенно начале их внепочечного развития. По мере роста побега и развертывания листьев, когда начинают преобладать процессы растяжения клеток, эта зависимость несколько ослабевает. По данным И. В. Серегина, А. Д. Кожевниковой (2006) ионы никеля подавляют дифференциацию клеток меристем, функционирующих в первые 10 дней развития листа, то есть в период начала формирования листовой пластинки.
5.2.2 Скорость развертывания листьев
В результате экспериментальных исследований в 2006 г. получено, что величина скорости развертывания листьев (рис. 5) изменяется в диапазоне: 9Ц80 суток (ПП5), 7Ц80 (ПП4), 9Ц20 (ПП3), 8Ц40 (ПП2), 7Ц40 (ПП1). Исследования показали, что величина скорости развертывания листьев в начале вегетации с конца мая до начала июня на фоновой площади и в зоне дефолиирующих лесов (ПП4, ПП3) высокая. Время, необходимое для развертывания каждого последующего листа в этот период в 2006 гг. составляет 9 суток на ПП5 и 4, 11 суток на ПП4, ПП3. (рис. 5). В техногенном редколесье и на техногенной пустоши в этот период скорость развертывания листьев намного меньше и составляет 25, 20 суток на ПП1 и ПП2 соответственно. Ингибирование развития листьев на этих двух площадках резко снижается к 20.06.2006, при этом скорость развития значительно возрастает. С середины июля 2006 г. скорость развертывания листьев уменьшается на всех площадках, а концу июля резко падает, особенно в фоне, что связано с замедлением развития листьев на побеге.

Рис. 5. Влияние техногенного загрязнения на скорость развертывания листьев
Betula czerepanovii в окрестностях комбината Североникель в 2006 г. (в сутках)
Наряду с естественной ритмикой развития листьев в условиях фона выявлено воздействие техногенного загрязнения на скорость их развертывания в ходе онтогенетического развития побега. В начале вегетационного периода в конце мая листья березы на площадках ПП1, ПП2 распускаются на 5 дней позже, затем в начале июня скорость их развертывания ингибирована и значительно ниже фоновых значений. В то же время в конце июля скорость развертывания листьев на этих площадках значительно выше, чем в условиях фона, где процесс образования новых листьев на побеге замедляется. Можно придти к заключению, что высокие концентрации никеля и меди изменяют ритмику развития листьев на побегах B. czerepanovii на техногенной пустоши и редколесье и период развертывания листьев на побеге удлиняется по сравнению с фоновыми значениями. Полученные результаты согласуются с данными Н. М. Казниной (2006) о том, что наибольшая степень ингибирования развития листьев наблюдается в начале вегетации, когда процесс их развертывания на побегах наиболее интенсивен.
5.2.3 Структура почек
Морфо-физиологические исследования терминальных почекпобеговB. czerepanovii перед началом вегетации выявили, что техногенное загрязнение не вызывает увеличения числа внешних почечных чешуй. Число зачаточных листьев в почках в конце мая как в 2005, так и в 2006 гг. незначительно уменьшается по мере приближения к комбинату Североникель (табл. 3), но эти различия не достоверны и находятся в пределах ошибки. Таким образом в зоне техногенного воздействия комбината не происходит ингибирования дифференциации апикальных меристем и процессов органогенеза в терминальных почках побегов B. сzerepanovii в период их внутрипочечного развития, которое длится с конца лета предшествующего года до конца мая текущего года.
Таблица 3
Влияние техногенного загрязнения на число почечных чешуй и зачаточных листьев в почках B. czerepanovii в окрестностях комбината Североникель в 2006 г. (шт.)
№ пробной площади |
Внешние чешуи N = 100 |
Зачаточные листья N = 100 |
Внутренние чешуи N = 100 |
|||
Mm |
Коэффициент вариации, % |
Mm |
Коэффициент вариации, % |
Mm |
Коэффициент вариации, % |
|
ПП1 |
7,60,9 |
11,7 |
5,60,6 |
10,1 |
9,30,5 |
5,8 |
ПП2 |
6,90,9 |
12,8 |
5,60,5 |
8,7 |
8,10,6 |
7 |
ПП3 |
6,60,7 |
10,2 |
6,10,7 |
11,7 |
7,70,8 |
10,5 |
ПП4 |
6,10,8 |
13,4 |
6,10,6 |
10,5 |
7,80,8 |
9,5 |
ПП5 |
6,20,9 |
13,7 |
6,60,8 |
12,2 |
7,90,7 |
9 |
Полученные данныесвидетельствуют о том, что число листьев, образуемых апикальными меристемами почек B. czerepanovii, детерминировано генетически, что проявляется в высокой устойчивости данного показателя даже в условиях острого хронического загрязнения среды тяжелыми металлами. По мнению Д. А. Сабинина (1963) образование листьев на побеге подчинено определенному морфогенетическому закону, их число является весьма устойчивым признаком, сохраняющимся в любых условиях. Закономерности, подтверждающие это положение, выявлены на многих видах растений (Bonaparte, 1975; Pearson, Coaldrake, 1983; Козлечков, 1986, 1988; Mohamed, Clark, 1988; Марковская и др., 1991; Харькина, Марковская, 1997; Василевская, 1991, 2007).
5.3 Флуктуирующая асимметрия листьев Betula czerepanovii
В ходе исследований B. сzerepanovii выявлено, что индекс флуктуирующей асимметрии листьев увеличивается по мере приближения к комбинату, что свидетельствует о возрастании нестабильности развития в популяции (табл. 4). В результате корреляционного анализаданных за 2006 г. получены высокие статистически значимые положительные корреляции (R > 0,9; р < 0,05) между содержанием тяжелых металлов в листьях и индексом ФА.
Высокие значения флуктуирующей асимметрии листьев B. сzerepanovii в зоне техногенного загрязнения комбината Североникель свидетельствуют о нарушении стабильности развития популяции и могут быть использованы как показатели раннего обнаружения стресса в условиях промышленного загрязнения. Полученные данные также подтвердили результаты аналогичных исследований популяций берез, проведенных ранее М. Козловым, Л. Зверевой в окрестностях г. Мончегорска (Kozlov et al., 2005) и г. Харьявалта (Финляндия) (Kozlov et al., 1996).
Таблица 4
Флуктуирующая асимметрия листьев B. czerepanovii
в окрестностях комбината Североникель (N = 100)
№ пробной площади |
ФА 2005 г. N = 100 |
ФА 2006 г. N = 100 |
ПП1 |
0,0970,003 |
0,0950,005 |
ПП2 |
0,0880,003 |
0,0870,004 |
ПП3 |
0,0820,003 |
0,0760,004 |
ПП4 |
0,0750,003 |
0,0490,003 |
ПП5 |
0,0330,002 |
0,0310,002 |
5.4 Репродуктивное развитие Betula czerepanovii в условиях техногенного загрязнения
5.4.1 Развитие соцветий Betula czerepanovii
В ходе исследований особенностей репродуктивного развития B. czerepanovii в зоне техногенного загрязнения комбината Североникель на площадках ПП3, ПП2, ПП1 отмечены отдельные особи с аномально большим количеством только мужских или только женских сережек. Доля таких деревьев на ПП1 составила 20%, на ПП2 Ч 30%, ПП3 Ч 40%. Установлено, что в течение трех вегетационных сезонов это были одни и те же деревья. На некоторых исследуемых особях сережки отсутствовали, на ПП1 доля таких деревьев составила 40%. На фоновой площадке деревьев с аномальным развитием не выявлено. В ходе исследований также получено, что наибольшее число мужских и женских сережек образуется на побегах B. czerepanovii на фоновой площадке. По градиенту загрязнения число мужских соцветий уменьшается, что особенно отчетливо выражено в вегетационные сезоны 2005 и 2007 гг. (табл. 5). Наибольшее ингибирование развития тычиночных соцветий отмечено на техногенной пустоши (ПП1).
Таблица 5
Влияние техногенного загрязнения на динамику мужских и женских сережек
B. czerepanovii (шт/побег) в окрестностях комбината Североникель (2005Ц2007 гг.)
№ пробной площади |
2005 год |
2006 год |
2007 год |
|||
> N = 50 |
+ N = 50 |
> N = 50 |
+ N = 50 |
> N = 50 |
+ N = 50 |
|
ПП1 |
1,320,47 |
1,680,56 |
1,440,50 |
1,120,43 |
1,760,56 |
1,260,47 |
ПП2 |
5,001,21 |
5,961,58 |
2,240,48 |
2,680,62 |
3,180,92 |
4,321,10 |
ПП3 |
3,960,70 |
5,660,96 |
1,700,46 |
2,260,65 |
2,580,73 |
4,500,99 |
ПП4 |
5,580,39 |
7,060,38 |
2,080,57 |
2,100,76 |
4,040,29 |
6,400,18 |
ПП5 |
5,900,30 |
8,160,90 |
2,780,71 |
3,320,74 |
4,500,02 |
7,240,10 |
Максимальное число женских сережек на протяжении трехлетнего мониторинга выявлено на фоновой площади. При этом на всех экспериментальных площадках, кроме техногенной пустоши, число женских соцветий больше, чем мужских (табл. 5). Наибольшее количество пестичных соцветий отмечено в вегетационные периоды 2005 и 2007 гг., по градиенту загрязнения число женских сережек на побеге уменьшается (табл. 5). На техногенной пустоши их количество минимальное во все вегетационные сезоны.
Согласно полученным нами данным в зонах хронического загрязнения ионами никеля и меди (ПП3, ПП2, ПП1) отмечается нестабильность репродуктивного развития B. czerepanovii, что выражается в усилении репродуктивной функции у ряда особей и резком увеличении на них числа пестичных или тычиночных соцветий, а также в нарушении репродуктивного развития у некоторых деревьев, которое проявляется в полном отсутствии на них мужских и женских сережек. В результате подсчета числа сережек на побегах мы получили, что в среднем число пестичных и тычиночных соцветий уменьшается по градиенту загрязнения.
Полученные нами результаты по уменьшению числа соцветий у березы по градиенту загрязнения не согласуются с данным группы исследователей университета Турку (Kozlov, Zvereva, 2004). Авторы отмечают, что большая часть взрослых деревьев березы в окрестностях комбината Североникель в 2002Ц2003 гг. не продуцировала ни мужских, ни женских сережек. Также по результатам исследований количество женских соцветий у берез достоверно увеличивалось вблизи источника загрязнения. При этом получено, что изменения в количестве мужских и женских сережек на исследуемых деревьях в течение двух лет наблюдений не связаны с уровнем техногенного загрязнения.
По мнению авторов (Kozlov, Zvereva, 2004) это обусловлено тем, что репродуктивные процессы в популяциях B. czerepanovii ибо обладают высокой устойчивостью к загрязнению, либо популяции, длительно подвергавшиеся воздействию сильного техногенного стресса, приобрели повышенную устойчивость к загрязнению вследствие элиминации в процессе естественного отбора наиболее чувствительных генотипов. Авторы высказали гипотезу, что такое увеличение репродуктивных возможностей в стрессовых условиях среды может рассматриваться как адаптационная реакция растительного организма на химическое загрязнение среды (Kozlov, Zvereva, 2004). Полученные нами результаты не свидетельствуют в пользу гипотезы об адаптации березы к химическому загрязнению среды, скорее мы наблюдаем стрессовое состояние популяцииB. czerepanovii, что проявляется в нарушении репродуктивного развития под воздействием высоких концентраций тяжелых металлов.
5.4.2 Жизнеспособность семян и всходов Betula czerepanovii
5.4.2.1 Грунтовая всхожесть семян
В целях изучения жизнеспособности семян, собранных с модельных деревьев B. czerepanovii на экспериментальных площадках, проведен лабораторный эксперимент. В результате получено, что прорастание семян B. czerepanovii отмечено во всех вариантах на 5-й день опыта (табл. 6). Массовые всходы отмечены на 7Ц10-й день, а максимальное количество проростков Ч на 20-й день. Наиболее интенсивное прорастание семяннаблюдалось в контроле, грунтовая всхожесть здесь достигла 69%. Значительное ингибирование прорастания отмечено в вариантах техногенной пустоши, редколесья и дефолиирующих лесов (ПП3). Грунтовая всхожесть B. czerepanovii в варианте техногенной пустоши на 5-й день была меньше, чем в контроле в 5 раз, на 7-й день в 3 раза, на 10-й, 15-й, 20-й в 2 раза. Близкие значения получены и в варианте техногенного редколесья. Энергия прорастания выше у семян B. czerepanovii в контроле (36%), во всех остальных вариантах значительно ниже. Семена B. сzerepanovii, собранные в зоне техногенного редколесья и техногенной пустоши имеют очень низкую энергию прорастания Ч 16% и 11% соответственно.
Низкая грунтовая всхожесть и энергия прорастания семян березы техногенной пустоши и техногенного редколесья свидетельствуют о снижении качества семян и жизнеспособности зародышей, нарушении процессов их эмбрионального развития в условиях высоких концентраций тяжелых металлов. Эти результаты подтверждают данные ряда авторов (Подзоров, 1965; Поповичев, 1980; Ставрова, 1990; Федорков, 1994; Бажина, 1998; Шихова, Козина, 2002), которые выявили, что в условиях техногенного загрязнения среды снижается качество и количество семян, жизнеспособность семенного потомства.
Таблица 6
Динамика грунтовой всхожести семян березы Betula czerepanovii (%)
Варианты опыта |
Доля проросших семян от количества высеянных семян |
|||||
3-й день |
5-й день |
7-й день |
10-й день |
15-й день |
20-й день |
|
1* |
0 |
4 |
11 |
22 |
31 |
34 |
2 |
1 |
7 |
16 |
26 |
34 |
38 |
3 |
1 |
12 |
24 |
32 |
45 |
49 |
4 |
3 |
14 |
25 |
41 |
52 |
55 |
5 |
5 |
19 |
36 |
52 |
69 |
69 |
*здесь и далее: 1 вариант Ч техногенная пустошь, 2 Ч техногенное редколесье, 3 Ч дефолиирующий лес (20 км), 4 Ч дефолиирующий лес (30 км), 5 Ч контроль (фоновая площадь).
5.4.2.2 Динамика роста Betula czerepanovii на ранних этапах онтогенеза
Изучение динамики роста проростков и ювенильных особей B. czerepanovii в ходе эксперимента показало, что во всех четырех вариантах опыта наблюдается ингибирование роста побега в длину по сравнению с контролем. Наиболее сильное ингибирование роста отмечено в варианте техногенной пустоши, на 7Ц10-й день опыта длина побега в 2,9Ц2,7 раза меньше, чем в контроле. В варианте техногенного редколесья данный показатель меньше контрольных значений в 2,8Ц2,5 раза, соответственно. К 15 дню ингибирующий эффект в этих двух вариантах снижается до 2,3 раза и остается на этом уровне до конца эксперимента. В варианте дефолиирующих лесов (ПП3) наиболее сильное ингибирование в 1,9 раза наблюдается на 7-й день, к 10 дню снижается до 1,5 раз и сохраняется до конца опыта. Результаты исследований показали, что семена березы, собранные на площадках, подверженных загрязнению тяжелыми металлами, не только имеют низкую грунтовую всхожесть и энергию прорастания, но впоследствии сеянцы березы имеют значительно более низкую динамику роста на ранних этапах онтогенеза, чем в контроле.

Рис.6. Длина побега ювенильных особей B. czerepanovii по вариантам опыта (мм)
(на 45-й день эксперимента)
Сведения о токсичности тяжелых металлов для растений, особенно на ранних этапах онтогенеза, немногочисленны и противоречивы. Показано, что негативное влияние тяжелых металлов на ростовые показатели связано, прежде всего, с их непосредственным действием на деление и растяжение клеток (Титов и др., 2007). Поэтому, рост главного побега можно рассматривать в качестве тестового показателя сформированности апекса побега зародыша семян и уровня дифференциации его клеток на ранних этапах онтогенеза растений.
5.4.2.3 Развитие Betula czerepanovii на ранних этапах онтогенеза
Исследования показателей развития проростков и ювенильных особей B. czerepanovii выявили, что растения значительно различались по вариантам опыта. Начало ювенильной фазы в контроле началось на 12-й день с начала эксперимента, в вариантах дефолиирующих лесов на 14-й день, в вариантах техногенной пустоши и редколесья на 17-й день. У особей вариантов техногенной пустоши и редколесья переход в ювенильное возрастное состояние произошел на пять дней позже, чем в контроле. Развитие настоящих листьев в этих двух вариантах значительно ингибировано в течение всей ювенильной фазы и к концу эксперимента их развернулось значительно меньше, чем в контроле: в 2,4 и 1,8 раза, соответственно. В варианте техногенной пустоши наблюдалось наибольшее число тератов и безлистных особей. У ряда особей отмечена недоразвитость и асимметричность настоящих листьев, при этом была недоразвита либо левая, либо правая половина листовой пластинки. Отмечена недоразвитость верхней части листовой пластинки, изменялась форма листа. Во всех вариантах опыта наблюдалось пожелтение настоящих листьев и ранняя дефолиация. Первые пожелтевшие листья появились в варианте техногенной пустоши на 20-й день эксперимента, техногенного редколесья на 22-й, дефолиирующих лесов Ч 30-й, в контроле Ч 35-й день эксперимента. Преждевременная дефолиация отмечена в вариантах техногенной пустоши и редколесья на 25-й день, это были единичные случаи, опадение листьев усилилось к концу эксперимента в период с 30 по 45-й день наблюдений.В вариантах дефолиирующих лесов опадение листьев у растений зафиксировано в период с 35 по 45-й день, оно не имело массового характера. В контроле первые опавшие листья отмечены только на 45-й день.
В ходе эксперимента в вариантах техногенной пустоши и редколесья отмечен высокий процент гибели растений. Большая часть проростков погибла в период с 20 по 30-й день в результате усыхания семядолей (табл. 7). В варианте техногенной пустоши на 45-е сутки эксперимента выжило только 65% ювенильных растений от их числа на 20-й день опыта (табл. 7). В варианте техногенного редколесья динамика развития особей несколько лучше, к концу эксперимента выжило 76% растений. В вариантах дефолиирующих лесов процент гибели небольшой, на 45-й день опыта осталось 84% и 87% особей, соответственно. В контроле отмечены лишь единичные случаи гибели растений,этот показатель не превысил 4%.
Таблица 7
Динамика численности проростков и ювенильных растений
Betula czerepanovii
Вариант опыта |
20-й день |
25-й день |
30-й день |
35-й день |
40-й день |
45-й день |
1* |
34 |
24 |
24 |
22 |
22 |
22 |
2 |
38 |
34 |
30 |
30 |
29 |
29 |
3 |
49 |
43 |
42 |
41 |
41 |
41 |
4 |
55 |
51 |
49 |
48 |
48 |
48 |
5 |
69 |
66 |
65 |
65 |
65 |
65 |
*1 вариант Ч техногенная пустошь, 2 Ч техногенное редколесье, 3 Ч дефолиирующий лес (20 км), 4 Ч дефолиирующий лес (30 км), 5 Ч контроль (фоновая зона).
В результате лабораторного эксперимента получено, что особи, выращенные из семянB. сzerepanovii техногенной пустоши и редколесья нежизнеспособны. В данных вариантах опыта у растений отмечено замедление развития, хлороз и пожелтение листьев, ранняя дефолиация, ингибирование роста побега, тератоморфные изменения листьев и побегов, гибель на ранних этапах онтогенетического развития, что связано с низкой жизнеспособностью семян, сформированных в условиях высокой концентрации никеля и меди в техногенной среде.
ЗАКЛЮЧЕНИЕ
На территории Евро-Арктического региона России, в условиях загрязнения среды медно-никелевым комбинатом Североникель (г. Мончегорск) изучено воздействие техногенного загрязнения на рост и развитие березы Черепанова. Получено, что содержание тяжелых металлов (Ni, Cu) в почве значительно увеличивается по градиенту загрязнения, эта зависимость имеет нелинейный характер. Изменяется состав и уменьшается число видов лихено-бриологической группы на площадках техногенной пустоши и редколесья. При этом ряд видов сосудистых растений выдерживает высокие концентрации тяжелых металлов и способствует формированию устойчивых сообществ. Такими видами являются: Betula czerepanovii, Salix borealis, Empetrum hermaphroditum и представители рода Vaccinium.
С увеличением содержания никеля и меди в почве по градиенту загрязнения происходит ингибирование роста побегов текущего года и размеров листьев в ходе онтогенеза, особенно в первые дни вегетации, в зонах техногенной пустоши и редколесья. Концентрации Ni и Cu в листьях B. czerepanovii многократно увеличиваются по градиенту загрязнения, при этом тяжелые металлы вызывают изменения мезоструктуры листа. Выявлена более значительная модификация палисадных клетокмезофилла B. czerepanovii, выражающаяся в их удлинении, уменьшении диаметра, увеличении индекса палисадности, усилении периклинальных делений. По мере приближения к комбинату Североникель увеличивается толщина листовой пластинки. Корреляционный анализ выявил статистически значимые (p < 0,05)сильныекорреляции между содержанием в листьях тяжёлых металлов и анатомическими параметрами.
Техногенное загрязнение ингибирует вегетативное развитие B. czerepanovii: число листьев на побегах текущего года, скорость их развертывания в первые дни вегетации в зонах техногенной пустоши и редколесья. Ингибирование развития листьев в начале их внепочечного роста свидетельствует о воздействии ионов никеля и меди на дифференциацию клеток в апексах терминальных почек именно в этот период. В то же время не выявлено влияния техногенного загрязнения на процессы органогенеза и число листьев в период внутрипочечного развития. По-видимому, число листьев, образуемых апикальными меристемами почек B. czerepanovii, детерминировано генетически, что проявляется в высокой устойчивости данного показателя даже в условиях острого хронического загрязнения. При расчетефлуктуирующей асимметрии листьев получены высокие значения индекса по градиенту загрязнения, что свидетельствуют о нарушении гомеостаза развития популяций B. сzerepanovii.
Более сложный характер реакций на воздействие техногенного загрязнения имеют показатели репродуктивного развития березы, произрастающей в окрестностях комбината. Выявлено, что по градиенту загрязнения возрастает доля деревьев с аномально высоким числом мужских или женских соцветий или полным их отсутствием. По мере возрастания техногенной нагрузки отмечается тенденция уменьшения числа мужских и женских соцветий. Эксперименты показали, что семена B. сzerepanovii с техногенной пустоши и редколесья имеют низкую грунтовую всхожесть и энергию прорастания, что свидетельствует о снижении качества семян и жизнеспособности зародышей, нарушении процессов их эмбрионального развития в условиях высоких концентраций тяжелых металлов. Растения, выращенные из семянB. сzerepanovii техногенной пустоши и редколесья нежизнеспособны, у них отмечено: замедление развития, хлороз и пожелтение листьев, ранняя дефолиация, тератоморфные изменения листьев и побегов, гибель на ранних этапах онтогенетического развития. В то же время, показатели роста и развития растений из семян B. сzerepanovii фоновой зоны значительно выше, что позволяет продолжить эксперименты по их возможному использованию для восстановления нарушенных территорий вокруг медно-никелевых производств.
ВЫВОДЫ
- Техногенное загрязнение ингибирует рост побегов и размеры листьев Betula czerepanovii в ходе онтогенеза, этот эффект наиболее выражен в первые дни вегетации в зонах техногенной пустоши и редколесья.
- Высокие концентрации никеля и меди в листьях B. czerepanovii техногенной пустоши и редколесья вызывают более значительную модификацию палисадных клетокмезофилла B. czerepanovii, чем губчатых, что выражается в их удлинении, уменьшении диаметра, увеличении индекса палисадности, усилении периклинальных делений.
- Высокое содержание никеля и меди в листьях B. czerepanovii техногенной пустоши и редколесья ингибируют их развитие и скорость развертывания в первые дни вегетации.
- Техногенное загрязнение не влияет на процессы органогенеза и число листьев березы в период внутрипочечного развития.
- Индекс флуктуирующей асимметриилистьевB. сzerepanovii увеличивается по градиенту загрязнения.
- Доля деревьев с аномальным плодоношением возрастает по градиенту загрязнения, при этом уменьшается число мужских и женских соцветий.
- Семена березы из зоны техногенного воздействия комбината Североникель имеют низкую грунтовую всхожесть и энергию прорастания в вариантах опыта техногенной пустоши и редколесья.
- Растения, выращенные из семянB. сzerepanovii техногенной пустоши и редколесья нежизнеспособны, для них характерно: замедление развития, хлороз и пожелтение листьев, ранняя дефолиация, ингибирование роста побега, тератоморфные изменения листьев и побегов, гибель на ранних этапах онтогенеза.
Статьи в рецензируемых журналах из списка ВАК:
- Василевская Н. В., Лукина Ю. М. Влияние техногенного загрязнения на динамику роста и мезоструктуру листьев Betula czerepanovii Orlova (Мурманская область) // Ученые записки Петрозаводского государственного университета. 2011. Серия Естеств. и техн. науки. Вып. 8. С. 14Ц18.
- укина Ю. М., Василевская Н. В. Воздействие промышленного загрязнения на строение листа Betula czerepanovii (Betulaceae) // Растительные ресурсы. 2012. Вып. 1. С. 53Ц59.
Публикации в других изданиях:
- Василевская Н. В., Лукина Ю. М. Флуктуирующая асимметрия листьев Betula pubescens ssp. subarctica Orlova, как интегральный показатель качества среды г. Мурманска // Ботанические проблемы регионального природопользования. Материалы чтений, посвященных памяти проф. Н. И. Антипова. Рязань, 2001. С.15Ц17.
- укина Ю. М., Василевская Н. В. Оценка стабильности развития Betula pubescens ssp. subarctica Orlova на территории г. Мурманска и его окрестностей // Естественнонаучные проблемы Арктического региона. Труды IV региональной научной студенческой конференции. Мурманск, 2003. С. 76Ц78.
- Василевская Н. В., Лукина Ю. М. Флуктуирующая асимметрия Betula czerepanovii Orlova как интегральный показатель качества среды г. Мурманска // Естественнонаучные проблемы Арктического региона. Тезисы докладов IV региональной научной студенческой конференции. Мурманск, 2003. С. 27Ц29.
- Василевская Н. В., Лукина Ю. М. Флуктуирующая асимметрия Pinus sylvestris L. как показатель стресса в условиях промышленного загрязнения Мурманской области // Экологические проблемы северных регионов и пути их решения. Часть 1. Материалы Межд. конференции. 31 августаЦ3 сентября 2004 г. Апатиты, 2004. С. 95Ц97.
- Василевская Н. В., Тумарова Ю. М. Оценка стабильности развития популяции Pinus sylvestris L. в условиях аэротехногенного загрязнения (Мурманская область) // Труды Карельского научного центра РАН. Вып. 7. Биогеография Карелии. Петрозаводск, 2005. С. 19Ц23.
- Василевская Н. В., Лукина Ю. М. Влияние аэротехногенного загрязнения на состояние популяции Betula czerepanovii Orlova в окрестностях комбината Североникель (Мурманская область) // Устойчивость экосистем и проблемы сохранения биоразнообразия на Севере. Материалы межд. конф. 26Ц30 августа 2006 г. Т. 1. Пленарные доклады. Флора и растительность. Кировск, 2006. С. 43Ц48.
- укина Ю. М. Флуктуирующая асимметрия листьев Betula czerepanovii Orlova как показатель качества среды в районе г. Мончегорска (Мурманская область) // Проблемы региональной экологии в условиях устойчивого развития. Материалы Всерос. научно-практической конф. с международным участием. Вып. 5, Ч. 1. Киров, 2007. С. 99Ц102.
- Василевская Н. В., Лукина Ю. М. Динамика роста и развития Betula czerepanovii в условиях промышленного загрязнения на Кольском полуострове // Экологическое образование и экологическая наука для устойчивого развития. Материалы 5 межд. научно-практической конференции. Архангельск, 2007. С. 208Ц212.
- укина Ю. М., Василевская Н. В. Влияние аэротехногенного загрязнения на репродуктивный потенциал Betula czerepanovii Orlova // Принципы и способы сохранения биоразнообразия. Материалы III Всерос. научной конф. Пущино, 2008. С. 261Ц262.
- укина Ю. М., Василевская Н. В. Репродуктивный потенциал Betula czerepanovii Orlova в условиях аэротехногенного загрязнения комбината Североникель // Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт. Мурманск, 2009. С. 157Ц160.
- укина Ю. М., Василевская Н. В. Влияние тяжелых металлов на фотосинтетический аппарат Betula czerepanovii Orlova // Физико-химические механизмы адаптации растений к антропогенному загрязнению в условиях Крайнего Севера. Тезисы докладов межд. научной конф. Апатиты, 2009. С. 199-201.
- Василевская Н. В., Лукина Ю. М. Состояние фотосинтетического аппарата Betula czerepanovii Orlova как показатель аэротехногенного загрязнения среды комбинатом Североникель (Мурманская область) // Промышленная ботаника: состояние и перспективы развития. Материалы VI межд. научной конф. Донецк, 2010. С. 89Ц90.
- укина Ю. М., Василевская Н. В. Влияние промышленных выбросов комбината Североникель на стабильность развития Betula czerepanovii Orlova // Экологические проблемы промышленных городов. Саратов, 2011. С. 244Ц246.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]