
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
ПОЧВООБИТАЮЩИЕ БЕСПОЗВОНОЧНЫЕ В ЭКОСИСТЕМАХ ХИБИНСКОГО ГОРНОГО МАССИВА
Автореферат кандидатской диссертации
На правах рукописи
ПОЖАРСКАЯ
Виктория Викторовна
ПОЧВООБИТАЮЩИЕ БЕСПОЗВОНОЧНЫЕ
В ЭКОСИСТЕМАХ ХИБИНСКОГО ГОРНОГО МАССИВА
03.02.04 - зоология
03.02.08 - экология
Автореферат
диссертации на соискание учёной степени кандидата
биологических наук
Петрозаводск - 2012
Работа выполнена в Федеральном государственном бюджетном учреждении науки Институте проблем промышленной экологии Севера Кольского научного центра Российской академии наук
Научные руководители |
кандидат биологических наук доцент Зенкова Ирина Викторовна кандидат биологических наук доцент Похилько Анна Антоновна |
Официальные оппоненты |
доктор биологических наук профессор Долгин Модест Михайлович |
кандидат биологических наук доцент Лябзина Светлана Николаевна |
аа
Ведущая организацияаа Федеральное государственное бюджетное учреждение науки Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Защита диссертации состоится 14 марта 2012 г. в 14 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете по адресу: 185910, Республика Карелия, Петрозаводск, пр. Ленина, 33; эколого-биологический факультет.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета. Электронная версия автореферата размещена на официальном сайтеа разослан л 11 февраля 2012 г.
Ученый секретарь диссертационного совета канд. биол. наукаа аа |
|
И.М. Дзюбук |

ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Горные экосистемы являются уникальными природными комплексами для исследования фундаментальных проблем биологии, экологииа и зоогеографии животных. В настоящее время не существует достоверной информации о разнообразии беспозвоночных животных горных районов ни по России в целом, ни по регионам. Предполагается, что экосистемы Северного Кавказа, Среднего и Южного Урала и юга Сибири в результате активного освоения представляют собой антропогенные модификации природных горных экосистем с богатым биоразнообразием. Горы менее освоенного Европейского Севера сохранили первозданный облик и природное богатство фауны и флоры.
Неоднозначны результаты сравнительных исследований почвенной фауны горных (интразональных) и равнинных (зональных) экосистем. По одним данным, структура населения, вертикальное размещение и численность животных существенно отличаются в зональных и горных почвах [Криволуцкий и др., 1997]. По другим источникам, почвы зональной тайги по составу и соотношению групп мезофауны идентичны горным аналогам [Грюнталь, 2009]. Разнообразие почвенной фауны предгорных и среднегорных ландшафтов Средней Сибири выше, чем на равнине [Стригнова, Порядина, 2005]. В высокогорных местообитаниях Урала, Алтая, Дальнего Востока влияние высотной поясности на беспозвоночных различно. Общей тенденцией является лишьа снижение видового богатства насекомых и смена таежных видов тундровыми с высотой.
Исследование фауны Хибин (Мурманская обл.) носило эпизодический характер. Первые сведения были обобщены В.Ю. Фридолиным [1934, 1936], который рассматривал Хибинскую горную страну как географический рефугиум, населённый сложным комплексом лесных, горных, арктоальпийских видов европейского и сибирского происхождения и редкими реликтовыми видами. Ученый описал около 1 тыс. видов насекомых. В дальнейшем были выполнены работы по жукам [Новиков, 1958; Россолимо, 1989, 1994] и дождевым червям [Рыбалов, 2006], показано разделение фауны жесткокрылых на лесной и тундровый ландшафтно-высотные комплексы и приуроченность тундровых видов жужелиц и стафилинид к высокогорным поясам. Обследование территории массива в рамках проекта по организации национального парка подтвердило зоогеографическую закономерность - обеднение энтомофауны Северной Евразии при продвижении с востока на запад [Эколого-экономическое обоснование Е, 1999].
Цель работысостояла в изучении своеобразия, структурно-функциональной организации и сезонной активности почвенной фауны Хибин с учетом факторов высотной поясности, экспозиции склонов и в сравнении с фауной зональных почв региона, изученных в достаточной степени.
Основными задачами были:
1. Исследование таксономического и трофического разнообразия и обилия почвенной фауны в высотно-поясном градиенте природных факторов.
2. Сравнение сообществ беспозвоночных в почвах разных гор Хибинского массива с целью выявления своеобразия или общих закономерностей их организации.
3. Сравнение разнообразия, структурно-функциональной организации и динамики сообществ беспозвоночных в горных и зональных почвах Мурманской области.
Научная новизна. Впервые детально исследована структурно-функциональная организация и динамика почвенной фауны в экосистемах Хибин. Проведено сравнение состава, численности, биомассы, трофической структуры и сезонной активности почвенной фауны в биоценозах горно-растительных поясов, склонов разной экспозиции и разных гор. Сформировано представление о типичной структуре почвенных зооценозов Хибин и степени экстремальности и/или благоприятности горных местообитаний. По результатам исследования таксономического и экологического разнообразия жесткокрылых сделан вывод о мягких климатических условиях в горных тундрах по сравнению с зональной кольской тундрой, а на основании видового и морфо-экологического разнообразия дождевых червей - заключение о высокой функциональной активности трофического комплекса сапрофагов в обогащенных биогенными элементами и высокогумусированных почвах Хибин по сравнению с почвами зональной тайги и тундры.
Обоснован вывод о фаунистическом своеобразии горных почв, обусловленном, с одной стороны, обеднением горной фауны видами, характерными для северо-таежных экосистем Мурманской области, с другой - наличием комплекса горных (аркто-альпийских и борео-монтанных) видов, нетипичных для зональной тайги. Установлены закономерности высотно-поясного распределения беспозвоночных совпадающие для ряда групп (жесткокрылые, многоножки, дождевые черви) с их широтно-зональным распространением по территории региона. Прослежена высотная смена таежных видов жуков тундровыми при сохранении во всех поясах преобладания по видовому составу и численности видов с лесной преференцией. Подтверждена характеристика субальпийского пояса как экотонного, переходного между лесным и тундровым поясами. Расширен региональный список доминирующих семейств жесткокрылых: стафилинид на 18 видов или 10%, жужелиц на 4 вида или 6%, щелкунов на 1 вид или 4%.
Теоретическая значимость. Сформировано представление о своеобразии почвенной фауны Хибинских гор, которое обусловлено их заполярным географическим положением, особыми физико-химическими и минералогическими характеристиками почв и специфическими условиями горного микроклимата. На основе фаунистических данных подтверждена принадлежность Хибинской горной провинции к северотаежной группе бореального класса типов поясности по классификации горных систем России [Урусевская, 2007]. Показано, что в почвах заполярных горных экосистем могут формироваться и функционировать разнообразные и продуктивные биотические сообщества, нетипичные для зональной тайги и тундры и обогащающие региональную фауну. Расширено представление о разнообразии и путях формирования фауны заполярных горных экосистем, что важно с точки зрения ее сохранения в условиях экстремальных природных факторов и антропогенного воздействия в промышленно развитых регионах Европейского Севера РФ.
Практическая значимость. Подготовлены к опубликованию дополненные и уточненные фаунистические списки жесткокрылых семейств Carabidae, Staphylinidae, Elateridae Хибинского горного массива и Мурманской области в целом. Жужелица блестящая Carabus nitens (Linnaeus, 1758), локальная популяция которой впервые обнаружена на территории региона в хибинских тундрах, рекомендована для включения в Красную книгу Мурманской области со статусом редкого (узколокального) вида.
Апробация работы. Результаты исследований озвучены на научных конференциях: Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт (Мурманск, 2009); Экология, эволюция и систематика животных (Рязань, 2009); Сбалансированное природопользование. Охрана природы Севера (Апатиты, 2010); Проблемы изучения лесов мерзлотной зоны (Якутск, 2011); III и IV всерос. конф. по лесному почвоведению (Петрозаводск, 2009; Апатиты, 2011); научн. форуме Ломоносов (Москва, 2010, 2011); межрег. совещ. энтомологов Сибири и Д. Востока (Новосибирск, 2010); XVI совещ. по почвенной зоологии (Ростов, 2011).
Публикации. По результатам диссертации опубликовано 14 работ, в том числе 3 статьи в рецензируемых журналах из списка ВАК РФ.
Структура и объем диссертации. Диссертация состоит из введения, 7 глав, выводов, 3 приложений и перечня литературы, включающего 124 источника, в т.ч. 25 на иностранных языках. Работа изложена на 145 страницах, содержит 38 рисунков, 58 таблиц.
Благодарности. Автор выражает искреннюю благодарность к.б.н. И.В. Зенковой и к.б.н. А.А. Похилько за руководство работой, идеи и постоянную помощь при подготовке диссертации; д.б.н. Г.А. Евдокимовой за возможность проведения исследований на базе Лаборатории экологии микроорганизмов ИППЭС КНЦ РАН, сотрудникам лаборатории за помощь в выполнении работы. Особую благодарность за идентификацию беспозвоночных, предоставленные данные и консультации выражаем д.б.н. Филиппову Б.Ю. (Архангельск), к.б.н. Колесниковой А.А. (Сыктывкар), к.б.н. Вершининой С.Д. (Екатеринбург), к.б.н. Рапопорт И.Б. (Нальчик), Барне А.Ж. (Москва), Шикову Е.В. (Тверь).
СОДЕРЖАНИЕ РАБОТЫ
ГЛАВА 1. ПРИРОДНЫЕ ОСОБЕННОСТИ
ХИБИНСКОГО ГОРНОГО МАССИВА
Приведены сведения об особенностях геологического строения, гидрологического режима, микроклимата, почв и биоты Хибинского горного массива. Освещена степень изученности почвенной фауны.
ГЛАВА 2. ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследована почвенная фауна трех гор, расположенных в юго-западной (горы Вудъяврчорр и Поачвумчорр) и юго-восточной (Ловчорр) частях Хибинского массива (табл.1). В период 2008-2011 гг. на склонах северной, северо-восточной, юго-восточной и южной экспозиции исследовано 14 биоценозов в пределах таежного, лесотундрового (субальпийского), тундрового (альпийского) горно-растительных поясов и в поясе полярной (арктической) пустыни на высоте от 300 до 1100 м над ур. м. За 6 полевых сезонов (весенний и осенний сезоны на каждой горе) обработано 200 почвенных проб и 290 ловушек (2900 лов./суток). Учтено более 12 тыс. экз. беспозвоночных: 5700 из образцов подстилки и 6600 экз. из ловушек
Зависимость фаунистических показателей от факторов поясности, высоты над ур. м. и экспозиции склонов оценивали по результатам дисперсионного анализа. Материалы по фауне горных почв сопоставляли с данными по лесным и тундровым почвам Мурманской области (рис.1), предоставленными сотрудниками ИППЭС КНЦ РАН. Сходство разнообразия беспозвоночных горных и зональных почв оценивали с помощью индекса Съеренсена и кластерного анализа (Statistica 7.0).
|
Рис. 1. Район исследования. Подзона северной тайги (66-67 с.ш.): ХГМ - Хибинский горный массив; Е1-Е5 - зональные ельники; С1-С5 - зональные сосняки. Зональная тундра (69 с.ш.): окрестности пос. Териберка и Дальние Зеленцы. |
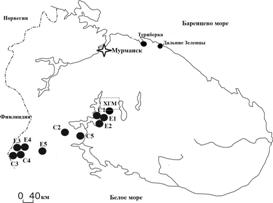
Табл. 1. Характеристика биоценозов, исследованных в пределах Хибин
Гора |
Экспозиция склона |
Высота м |
Биоценоз |
Условное обозначение |
Количество |
|
проб |
ов./сут. |
|||||
овчорр |
1093 |
Арктическая пустыня |
-ГП-ю |
60 |
800 |
|
Южная |
735 |
Тундра кустарничковая |
-ТК-ю |
|||
Северная |
452 |
Березовое криволесье |
-БК-с |
|||
Поачвумчорр |
Северная |
410 |
Тундра кустарничковая |
П-ТК-с |
72 |
1200 |
Юго-восточная |
400 |
Березовое криволесье |
П-БК-юв |
|||
Южная |
400 |
Березовое криволесье |
П-БК-ю |
|||
390 |
Еловое редколесье |
П-ЕР-ю |
||||
Вудъяврчорр |
Северо-восточная |
478 |
Тундра лиш.-куст. |
В-ТЛК-св |
66 |
900 |
370 |
Тундра кустарничковая |
В-ТК-св |
||||
366 |
Березовое криволесье |
В-БК-св |
||||
357 |
Еловое редколесье |
В-ЕР-св |
||||
Южная |
390 |
Березовое криволесье |
В-БК-ю |
|||
333 |
Тундра кустарничковая |
В-ТК-ю |
||||
324 |
Тундра ерниковая |
В-ЕРН-ю |
||||
ГЛАВА 3. ПОЧВЕННАЯ ФАУНА ГОРЫ ВУДЪЯВРЧОРР
3.1. Склон северо-восточной экспозиции
3.1.1. Горно-таежный пояс (еловое редколесье разнотравно-зеленомошное)
Выявлено 17 таксонов беспозвоночных животных (14 весной и 9 осенью), из них 7 присутствовали в подстилках в оба сезона: энхитреиды, моллюски, пауки, мягкотелки, жужелицы, стафилиниды, личинки двукрылых. Идентифицировано 20 видов жуков (стафилинид 12, жужелиц 5, щелкунов 3), 2 вида дождевых червей, 3 вида подстилочных моллюсков: Discus ruderatus (Ferussac, 1821), Nesovitrea petronella (L. Pfeiffer, 1853), Arion subfuscus (Draparnaud, 1805).
Средняя за два сезона сбора численность беспозвоночных составила 430 экз./м2, средняя биомасса (сырая) - 2.48 г/м2. Весенние и осенние значения таксономического состава, численности и биомассы отражены в табл. 2. Высокой плотности (100-300 экз./м2) достигали жуки и беспозвоночные с коротким периодом развития в почве - энхитреиды и личинки двукрылых. Весной доминантами с долей 10% и более были энхитреиды (56) и двукрылые (22). Осенью преобладали двукрылые (35%), мягкотелки (17), стафилиниды (21), энхитреиды (12) и пауки 5%.
По биомассе преобладали сапрофаги с общей долей более 90%: весной моллюски - 26, дождевые черви - 23, энхитреиды - 19, личинки двукрылых - 15, проволочники - 10%, а осенью - слизни (92%).
Трофическая структура включала 6 таксонов зоофагов и по 5 групп сапрофагов и насекомых-фитофагов.
3.1.2. Субальпийский пояс (березовое криволесье кустарничковое)
Выявлены те же 17 таксонов беспозвоночных, что и в горно-таежном поясе. В отличие от горного ельника в лиственной подстилке березняка в середине сентября сохранялись крупные сапрофаги - дождевые черви и проволочники, но отсутствовали насекомые-фитофаги. Определено 17 видов жуков (стафилиниды 11, жужелицы и щелкуны по 3), 3 вида люмбрицид, 2 вида моллюсков: N. petronella и Zoogenetes harpa (Say, 1824).
Табл 2. Структура почвенной фауны северо-восточного склона г. Вудъяврчорр
Показатели |
Число таксонов |
Численность, экз./м2 |
Биомасса, мг/м2 |
|||
Весна |
Осень |
Весна |
Осень |
Весна |
Осень |
|
Горно-таежный пояс |
||||||
Всего |
14 |
9 |
512 |
290 |
2681 |
2197 |
Сапрофаги, С |
5 |
3 |
447 |
145 |
2473 |
2023 |
Зоофаги, З |
6 |
4 |
43 |
135 |
114 |
172 |
Фитофаги, Ф |
3 |
2 |
25 |
10 |
94 |
2 |
Соотношение С:З:Ф, % |
1.7:2:1 |
1.5:2:1 |
87:8:5 |
50:47:3 |
92:4:4 |
92:8:<1 |
Субальпийский пояс |
||||||
Всего |
15 |
7 |
713 |
454 |
13189 |
718 |
Сапрофаги, С |
5 |
4 |
632 |
325 |
12952 |
550 |
Зоофаги, З |
6 |
3 |
67 |
129 |
133 |
168 |
Фитофаги, Ф |
4 |
0 |
14 |
0 |
104 |
0 |
С:З:Ф, % |
1.2:1.5:1 |
4:3:0 |
89:9:2 |
70:30:0 |
98:1:1 |
76:23:0 |
Тундра кустарничковая |
||||||
Всего |
13 |
8 |
426 |
296 |
2572 |
638 |
Сапрофаги, С |
5 |
2 |
164 |
104 |
1959 |
58 |
Зоофаги, З |
5 |
5 |
89 |
179 |
376 |
578 |
Фитофаги, Ф |
3 |
1 |
173 |
13 |
237 |
2 |
С:З:Ф, % |
1.7:1.7:1 |
2:5:1 |
38:22:40 |
35:61:4 |
76:15:9 |
9:90:<1 |
Тундра лишайниково-кустарничковая |
||||||
Всего |
11 |
10 |
177 |
175 |
198 |
221 |
Сапрофаги, С |
4 |
3 |
138 |
90 |
35 |
58 |
Зоофаги, З |
5 |
4 |
29 |
70 |
148 |
159 |
Фитофаги, Ф |
2 |
3 |
10 |
15 |
15 |
4 |
С:З:Ф, % |
2:2.5:1 |
1:1.3:1 |
79:16:5 |
51:40:9 |
17:75:8 |
26:72:2 |
Численность в оба сезона была в 1.5 раза выше, чем в горном ельнике, составив в среднем ~600 экз./м2 (табл. 2). Весной, как и в ельнике, преобладали двукрылые и энхитреиды с плотностью более чем по 250 экз./м2 и долей в составе фауны 36-38%. Высокой численности (до 70 экз./м2 или 10%) достигали дождевые черви и жесткокрылые, а среди жуков - щелкуны (до 30 экз./м2), жужелицы и стафилиниды (по 15 экз./м2). Осенью доминировали только личинки двукрылых (63%) и стафилиниды (25%) с плотностью более 100 экз./м2. Биомасса весной достигала значений, высоких как для горных почв, так и зональных лесных подзолов (более 13 г/м2) при супердоминировании ключевой группы сапрофагов - дождевых червей (90%). Осенью биомасса сократилась до 1 г/м2, её формировали личинки двукрылых (64%), жужелицы (16), дождевые черви (10) и стафилиниды (7). В лиственном лесу в оба сезона сохранялось преобладание сапрофагов (дождевых червей, энхитреид, личинок двукрылых и щелкунов). В сравнении с ельником обилие сапрофагов было выше, фитофагов - ниже, хищников - сопоставимо.
3.1.3. Альпийский (горно-тундровый) пояс
3.1.3.1. Тундра кустарничковая
Тундра кустарничковая по составу таксонов не уступала горно-лесным поясам, а в июне отличалась большим разнообразием видов дождевых червей и жужелиц. Из 17 таксонов в оба сезона встречены 6: сапрофильные личинки щелкунов и двукрылых и активно мигрирующие хищники - многоножки, пауки, жужелицы и стафилиниды. Осенью таксономический состав был обеднен тонкопокровными моллюсками, червями (нематодами, энхитреидами, люмбрицидами) и насекомыми-фитофагами (чешуекрылыми, щитовками, долгоносиками). Выявлено 9 видов жесткокрылых (стафилинид 1, жужелиц 6, щелкунов 2), 4 вида дождевых червей, 2 вида моллюсков (Z. harpa и слизень A. subfuscus).
Численность и биомасса были сопоставимы с горно-таежным поясом и характеризовалась той же динамикой: снижались к осени (табл. 2). Сходной в этих поясах была и осенняя структура фауны. Доминировали двукрылые (100 экз./м2 или 30%) и жуки (150 экз./м2, 52%), в т.ч. мягкотелки (30%), стафилиниды и жужелицы, а также пауки (7-11%). По биомассе преобладали хищники - пауки (54%) и жуки (33): жужелицы (14), мягкотелки (11), стафилиниды (5%). Весной массово размножались фито- (микофаги) щитовки, типичные для тундровых ландшафтов: 170 экз./м2 (40%). В подстилках горно-лесных поясов фитофаги были единичны, независимо от сезона, а доминирующие в них группы в альпийском поясе были малочисленнее щитовок: двукрылые в 2 раза, пауки в 2.8, энхитреиды в 4.5, жесткокрылые в 5 раз, доля каждой из них не превысила 19%. По биомассе весной преобладали дождевые черви и моллюски (суммарно 70%), субдоминировали жуки и пауки (7-8%). Осенью преобладали хищники: пауки (54%), жужелицы и мягкотелки (11-14), многоножки и стафилиниды (5-7%). Осенью состав сапро- и фитофагов обеднялся, их численность и биомасса сокращались, показатели обилия зоофагов возрастали.
3.1.3.2. Тундра лишайниково-кустарничковая
Отличалась наименьшими в высотном ряду значениями разнообразия, численности и биомассы фауны и незначительным сезонным варьированием этих показателей (табл. 2). Из 13 таксонов в оба сезона встречались 8: сапрофильные энхитреиды, личинки двукрылых и щелкунов, хищные многоножки, пауки, стафилиниды, жужелицы, фитофаги щитовки. Идентифицировано 2 вида жуков, не встреченных в ниже лежащих поясах: голарктический аркто-альпийский вид жужелиц Curtonotus alpinus (Paykull, 1790) и трансевразиатский аркто-борео-монтанный щелкун тундровый Hypnoidus rivularius (Gyllenhal, 1808). Дождевые черви и моллюски не выявлены.
Численность и биомасса отличались минимальными по склону значениями (~180 экз./м2, ~200 мг/м2) и слабым сезонным варьированием. В оба сезона доминировали двукрылые: 68% от общей численности фауны весной и 37% осенью. Остальные таксоны были единичны. По массе весной преобладали жужелицы (40%) и пауки (8), осенью наряду с жужелицами и пауками (по 30%) в число доминантов входили многоножки и энхитреиды. Трофическая структура включала 5 таксонов зоофагов, 4 таксона сапрофагов и 3 группы фитофагов. В оба сезона численно преобладали сапрофаги (за счет двукрылых), по биомассе - зоофаги (<70%). Как и в ниже лежащих поясах осенью обилие сапро- и фитофагов снижалось, хищников - возрастало.
3.2. Склон южной экспозиции
3.2.1. Субальпийский пояс (березовое криволесье травянистое)
Осенью выявлены 9 таксонов, как и на северо-восточном склоне. Общими для березняков двух склонов были 5 таксонов: энхитреиды, дождевые черви, двукрылые, пауки и стафилиниды. Березняк на южном склоне отличался присутствием мягкотелок, многоножек геофилид и литобиид, жуков-долгоносиков. Осенняя численность была в 2 раза ниже по сравнению с северо-восточным склоном (220 экз./м2 против 460 экз./м2) из-за достоверно меньшей плотности двукрылых (в 40 раз: 7 экз./м2 против 300) и стафилинид (в 16 раз: 7 экз./м2 против 120). Прочие сапрофаги (энхитреиды, дождевые черви) и хищники (многоножки, пауки, мягкотелки) были в 3-16 раз обильнее на южном склоне. Доминировали энхитреиды (60%, 130 экз./м2), дождевые черви (16%, 36 экз./м2), пауки и многоножки-костянки (5-7%, 11-14 экз./м2). Биомасса достигала 11 г/м2 и была максимальной среди ее осенних значений в других биоценозах г. Вудъяврчорр. Супердоминировали люмбрициды (95% всей массы). В трофической структуре преобладали сапрофаги (эхитреиды, люмбрициды, двукрылые) (табл. 3).
3.2.2. Альпийский пояс (тундра кустарничковая)
По сравнению с субальпийским поясом в альпийском состав фауны был разнообразнее (13 таксонов), численность в 4 раза выше (~800 экз./м2), биомасса в 2 раза меньше (5 г/м2) (табл. 3). Трофические группы зоофагов (пауки, многоножки-костянки, мягкотелки, жужелицы и стафилиниды) и сапрофагов (нематоды, энхитреидами, дождевые черви, личинки двукрылых и щелкуны) включали по 5 таксонов, фитофаги - 3 (тли, перепончатокрылые и долгоносики). Преобладали энхитреиды (540 экз./м2 или 68%), личинки двукрылых и многоножки (по 7%). Распределение биомассы было равномерным: энхитреиды (31%), дождевые черви (27), щелкуны (17), многоножки (12).
3.2.3. Тундра ерниковая
Участок ерниковой тундры отличался наиболее низкими значениями разнообразия и обилия беспозвоночных среди биоценозов южного склона г. Вудъяврчорр. Из 8 таксонов 3 относились к сапрофагам (энхитреиды, личинки двукрылых и щелкунов), 4 к зоофагам (мягкотелки, жужелицы и стафилиниды, муравьи) и только 1 (клопы) к фитофагам. Крупные сапрофаги (дождевые черви и моллюски), как и активные хищники (пауки и многоножки) не выявлены.
Численность и биомасса были в 2 раза ниже, чем в соседней тундре кустарничковой (табл. 3), но выше чем в субальпийском поясе из-за обилия основных доминантов - энхитреид и личинок двукрылых (75-110 экз./м2 или 26-28%). Высокой численности достигали стафилиниды (42 экз./м2) и жужелицы (17). Остальные представители фауны были единичными. По биомассе преобладали личинки щелкунов (36), двукрылых (29), жужелицы (17) и энхитреиды (12%).
Табл. 3. Структура почвенной фауны южного склона г. Вудъяврчорр (осень)
Показатели |
Число таксонов |
Численность, экз./м2 |
Биомасса, мг/м2 |
Субальпийский пояс |
|||
Всего |
9 |
218 |
10957 |
Сапрофаги, С |
3 |
171 |
10561 |
Зоофаги, З |
5 |
43 |
304 |
Фитофаги, Ф |
1 |
4 |
92 |
С:З:Ф, % |
3:5:1 |
79:19:2 |
96:3:1 |
Альпийский пояс |
|||
Всего |
13 |
795 |
4835 |
Сапрофаги, С |
5 |
660 |
3913 |
Зоофаги, З |
5 |
100 |
662 |
Фитофаги, Ф |
3 |
35 |
260 |
С:З:Ф, % |
1.7:1.7:1 |
83:13:4 |
81:14:5 |
Ерниковая тундра |
|||
Всего |
8 |
392 |
2207 |
Сапрофаги, С |
3 |
292 |
1707 |
Зоофаги, З |
4 |
92 |
499 |
Фитофаги, Ф |
1 |
8 |
1 |
С:З:Ф, % |
3:4:1 |
75:23:2 |
77:22:1 |
ГЛАВА 4. ПОЧВЕННАЯ ФАУНА ГОРЫ ПОАЧВУМЧОРР
4.1. Склон южной экспозиции
4.1.1. Горно-таежный пояс
(ельник-зеленомошник травянисто-мохово-кустарничковый)
Выявлено 20 таксонов беспозвоночных животных, 15 встречались в оба сезона: сапрофильные нематоды, энхитреиды, дождевые черви, моллюски, личинки двукрылых и щелкунов, хищные мягкотелки, стафилиниды, многоножки, пауки, сенокосцы, муравьи, сосущие фитофаги - клопы, трипсы, щитовки. Осенью не обнаружены жужелицы, чешуекрылые и цикадки, но выявлены жуки листоеды и мертвоеды Necrophorus vespilloides (Herbst, 1783). Определено 2 вида дождевых червей, 21 вид стафилинид, 1 вид жужелиц и 3 вида щелкунов.
Численность незначительно варьировала по сезонам, составив в среднем 560 экз./м2. В оба сезона доминировали личинки двукрылых (24-30%), только они достигали численности более 100 экз./м2. Осенью были обильны пауки (120 экз./м2, 20%). Доля щелкунов и стафилинид в оба сезона составляла 10-15%, энхитреиды - по 5%. Средние значения биомассы составляли 4 г/м2. Доминантный комплекс формировали в основном сапрофаги: весной дождевые черви (47%), личинки щелкунов и двукрылых (по 15%), осенью - моллюски (47%), дождевые черви (23), личинки двукрылых (18) и щелкунов (6%). Трофическая структура была сбалансирована: 7 таксонов зоофагов, по 6 фито- и сапрофагов при доминировании последних по численности и биомассе (табл. 4).
Табл. 4. Структура почвенной фауны в биоценозах г. Поачвумчорр
Показатели |
Число таксонов |
Численность, экз./м2 |
Биомасса, мг/м2 |
|||
Весна |
Осень |
Весна |
Осень |
Весна |
Осень |
|
Горно-таежный пояс (южная экспозиция) |
||||||
Всего |
17 |
16 |
502 |
602 |
1724 |
5806 |
Сапрофаги, С |
5 |
6 |
288 |
325 |
1402 |
5448 |
Зоофаги, З |
7 |
6 |
154 |
248 |
233 |
342 |
Фитофаги, Ф |
5 |
4 |
60 |
29 |
89 |
16 |
С:З:Ф, % |
1:1.4:1 |
1.5:1.5:1 |
57:31:12 |
54:41:5 |
79:13:5 |
93:6:1 |
Субальпийский пояс (южная экспозиция) |
||||||
Всего |
14 |
18 |
486 |
695 |
10084 |
12287 |
Сапрофаги, С |
5 |
5 |
286 |
446 |
8830 |
11793 |
Зоофаги, З |
6 |
7 |
150 |
211 |
1165 |
410 |
Фитофаги, Ф |
3 |
6 |
50 |
38 |
89 |
84 |
С:З:Ф, % |
1.7:2:1 |
1:1.4:1.2 |
59:31:10 |
64:30:6 |
88:11:1 |
96:3:1 |
Субальпийский пояс (юго-восточная экспозиция) |
||||||
Всего |
13 |
14 |
332 |
706 |
2259 |
7987 |
Сапрофаги, С |
6 |
5 |
272 |
478 |
2102 |
7710 |
Зоофаги, З |
6 |
6 |
58 |
208 |
156 |
273 |
Фитофаги, Ф |
1 |
3 |
2 |
20 |
1 |
4 |
С:З:Ф, % |
6:6:1 |
1.7:2:1 |
81:18:1 |
68:29:3 |
92:7:1 |
96:3:1 |
Альпийский пояс (северная экспозиция) |
||||||
Всего |
11 |
13 |
382 |
760 |
1362 |
953 |
Сапрофаги, С |
3 |
3 |
244 |
589 |
353 |
367 |
Зоофаги, З |
6 |
6 |
116 |
163 |
796 |
352 |
Фитофаги, Ф |
2 |
4 |
22 |
8 |
213 |
234 |
С:З:Ф, % |
1.5:3:1 |
1:2:1.3 |
64:30:6 |
78:21:1 |
26:58:16 |
39:37:24 |
4.1.2. Субальпийский пояс (березовое криволесье травянистое)
Выявлено 20 таксонов беспозвоночных, из них 14 - в оба сезона: сапрофильные нематоды, энхитреиды, дождевые черви, моллюски, личинки двукрылых и щелкунов, хищные многоножки-костянки, пауки, мягкотелки, жужелицы, стафилиниды, муравьи, фитофаги клопы и долгоносики. Осенью отсутствовали чешуекрылые, но разнообразие было выше за счет многоножек-землянок и фитофагов - цикадок, трипсов, равнокрылых и листоедов. Разнообразие стафилинид (23 вида), жужелиц (3), щелкунов (6) и дождевых червей (5 видов) было выше, чем в соседнем горно-таежном поясе, а для стафилинид и дождевых червей - максимальным среди 14 исследованных горных биоценозов.
Средняя численность составила ~600 экз./м2, при незначительном сезонном варьировании (табл. 4). К осени увеличилось число групп с численностью более 100 экз./м2 (личинки двукрылых и энхитреиды), весной столь многочисленными были только личинки щелкунов. Средняя биомасса составила более 11 г/м2 за счет дождевых червей (80% весной и 62% осенью). В трофической структуре по всем показателям преобладали сапрофаги. В комплексе зоофагов весной доминировали стафилиниды (32%), многоножки-костянки и пауки (по 21), по биомассе - муравьи (65), многоножки и жужелицы (10-14%). Фитофаги и зоофаги были разнообразнее осенью.
4.2. Склон юго-восточной экспозиции
4.2.1. Субальпийский пояс (березняк мохово-травянистый)
Выявлено 16 таксонов, 11 в оба сезона: нематоды, энхитреиды, дождевые черви, личинки двукрылых и щелкунов, многоножки-костянки, пауки, муравьи, мягкотелки, жужелицы, и стафилиниды. Весной не выявлены моллюски и фитофаги - клопы, трипсы, цикадки, в сентябре не обнаружены долгоносики. По сравнению с березняком травянистым на южном склоне разнообразие жужелиц увеличилось до 8 видов, щелкунов уменьшилось до 2, дождевых червей - до 4 видов. Состав стафилинид по-прежнему насчитывал 23 вида, общими для двух березняков были 20 видов (коэф. Съеренсена 47%), общими для трех лесных биоценозов горы - 12 видов (сходство 35%), что отражает различия почвенных свойств в этих биоценозах.
Средняя численность составила ~570 экз./м2. В оба сезона доминировали личинки двукрылых (по 38%), только они достигали численности более 100 экз./м2. Средняя биомасса была ~5 г/м2. В оба сезона доминировали только сапрофаги: весной - дождевые черви (66%), проволочники (19) и двукрылые (6), осенью - дождевые черви (71), двукрылые (11), энхитреиды и моллюски (по 6). В трофической структуре в оба сезона преобладали сапрофаги: 68-81% от общей численности и 92-96% биомассы. Среди зоофагов наблюдалась смена доминирования: весной доминировали пауки, стафилиниды и многоножки; осенью - стафилиниды, пауки и мягкотелки. В трофической группе фитофагов наблюдалась полная смена таксонов и расширение состава к осени (табл. 4).
4.3. Склон северной экспозиции
4.3.1. Альпийский пояс (тундра кустарничково-мохово-лишайниковая)
Выявлено 16 групп беспозвоночных, в оба сезона встречались 10: нематоды, энхитреиды, личинки двукрылых и щелкунов, многоножки-костянки, пауки, мягкотелки, жужелицы, стафилиниды и долгоносики. Разнообразие дождевых червей (1 вид - Dendrobaena octaedra (Savigny, 1826)) и стафилинид (12 видов) было минимальным в пределах горы, жужелиц (10) и щелкунов (6 видов), напротив, максимальным. Только в данном биоценозе выявлено 17 экз. имаго жужелицы Carabus nitens (Linnaeus, 1758) - европейского вида, редкого на территории европейской части России и включенного в Красные книги Рязанской, Калужской, Тверской, Московской, Владимирской областей и Республик Карелия и Коми как редкий, уязвимый, нуждающийся в охране вид.
Средняя численность была ~600 экз./м2, при двукратном увеличении осенью. В оба сезона доминировали личинки двукрылых (53-69%), пауки и стафилиниды (6-9). Также осенью доминантный комплекс включал многоножек и мягкотелок, а весной - личинок щелкунов (7-9%). Средняя биомасса составила ~1.1 г/м2. В оба сезона доминировали двукрылые (18-30%), мягкотелки (10-18) и проволочники (7-9). Весной - еще и пауки (30%), многоножки и долгоносики (по 14), а осенью - пилильщики-ткачи (24) и жужелицы (7). В трофической структуре по составу и биомассе преобладали зоофаги (табл. 4).
ГЛАВА 5. ПОЧВЕННАЯ ФАУНА ГОРЫ ЛОВЧОРР
5.1. Субальпийский пояс (березовое криволесье кустарничковое)
Выявлено 20 таксонов беспозвоночных,а 11 групп в оба сезона: энхитреиды, двукрылые, щелкуны, многоножки, пауки, мягкотелки, жужелицы, стафилиниды, долгоносики, щитовки и клопы. Крупные сапрофаги - дождевые черви и моллюски, фитофаги - гусеницы чешуекрылых, тли, листоеды, пантофаги - муравьи обнаружены только осенью, нематоды, трипсы и бриофаги - пилюльщики - только весной. Отмечено 3 вида моллюсков - обитателей лиственной подстилки (Euconulus fulvus (O.F. Muller, 1774), D. ruderatus, Z. harpa), 12 видов стафилинид, 6 видов щелкунов и наибольшее из среди горных биоценозов разнообразие жужелиц (14 видов).
Средняя численность составила 370 экз./м2. В оба сезона преобладали двукрылые (36-62%) и жесткокрылые (20-46) с численностью более чем по 100 экз./м2. Весной доли более 10% достигали щелкуны, 5-10% - стафилиниды, мягкотелки, долгоносики и гумификаторы-энхитреиды. Биомасса составила ~860 мг/м2. Весной преобладали жесткокрылые: сапрофилиные личинки щелкунов (36%), бриофаги - пилюльщики (17), фитофаги - долгоносики (12). Субдоминировали личинки двукрылых (12%), мягкотелки (8) и энхитреиды (5). Осенью преобладали крупные сапрофаги: дождевые черви (30), моллюски и щелкуны (9-11%), а также личинки двукрылых (20). Трофическая структура включала сапро- и зоофагов - по 6 групп, фитофагов - 7 групп. В сезонном аспекте наиболее стабильным был комплекс зоофагов, наиболее изменчивым - состав фитофагов. В оба сезона >70% численности и биомассы составляли сапрофагов (табл. 5).
5.2. Альпийский пояс (тундра лишайниково-кустарничково-моховая)
Выявлено 17 таксонов, из которых осенью отсутствовали дождевые черви и насекомые-фитофаги: клопы, щитовки, гусеницы чешуекрылых. В оба сезона не встречены трипсы и тли, характерные для субальпийского пояса этой горы. Моллюски были представлены только слизнями A. subfuscus. Выявлено 23 вида жуков: стафилинид 13, жужелиц 8, щелкунов 2. Общими с субальпийским поясом были 6 видов стафилинид, 4 вида жужелиц и 2 вида щелкунов.
Средняя численность (588 экз./м2) была в 1.5 раза выше, чем в субальпийском поясе (374 экз./м2) за счет массового размножения типичных для тундровых экосистем насекомых-мицетофагов - щитовок. Их численность весной достигала 290 экз./м2, доля в составе фауны - 42%. Высокую плотность (134 экз./м2 или 20%) имели энхитреиды, также типичные для тундровых почв. Доминантный комплекс дополняли двукрылые (11), стафилиниды и многоножки (5-6). Осенью возросла плотность двукрылых (в 5 раз, до 380 экз./м2 или 77%) и стафилинид (в 1.5 раза, до 60 экз./м2 или 12%). Численность каждой из остальных групп не превысила 10 экз./м2, доля - 3%. Биомасса, как и численность, в весенний сезон была выше, чем в субальпийском поясе из-за присутствия дождевых червей и более высокой массы энхитреид (в 4 раза) и многоножек (в 19 раз). Осенью наиболее значительно (в десятки раз) сократилась биомасса жесткокрылых разных трофических групп, менее резкое (в 4 раза) - у энхитреид и многоножек. У двукрылых, щелкунов, стафилинид, моллюсков и пауков, напротив, биомасса возрастала в 1.5-9 раз.
Табл. 5. Структура почвенной фауны в биоценозах г. Ловчорр
Показатели |
Число таксонов |
Численность, экз./м2 |
Биомасса, мг/м2 |
|||
Весна |
Осень |
Весна |
Осень |
Весна |
Осень |
|
Субальпийский пояс (северная экспозиция) |
||||||
Всего |
13 |
17 |
297 |
449 |
766 |
928 |
Сапрофаги, С |
4 |
5 |
216 |
325 |
548 |
662 |
Зоофаги, З |
5 |
6 |
59 |
110 |
120 |
167 |
Фитофаги, Ф |
4 |
6 |
22 |
14 |
98 |
99 |
С:З:Ф, % |
1:1.3:1 |
1:1.2:1.2 |
73:20:7 |
72:25:3 |
70:16:14 |
71:11:18 |
Альпийский пояс (южная экспозиция) |
||||||
Всего |
16 |
12 |
681 |
497 |
1769 |
568 |
Сапрофаги, С |
6 |
5 |
261 |
410 |
1163 |
353 |
Зоофаги, З |
5 |
5 |
103 |
82 |
307 |
197 |
Фитофаги, Ф |
5 |
2 |
317 |
5 |
299 |
18 |
С:З:Ф, % |
1.2:1:1 |
2.5:2.5:1 |
38:15:47 |
82:17:1 |
66:17:17 |
62:35:3 |
Пояс полярной (арктической) пустыни |
||||||
Всего |
9 |
12 |
217 |
133 |
340 |
514 |
Сапрофаги, С |
3 |
3 |
167 |
69 |
137 |
352 |
Зоофаги, З |
4 |
6 |
45 |
58 |
197 |
158 |
Фитофаги, Ф |
2 |
3 |
5 |
6 |
6 |
4 |
С:З:Ф, % |
1.5:2:1 |
1:2:1 |
77:21:2 |
52:43:5 |
40:58:2 |
68:31:1 |
Структура доминирования сообщества по биомассе в оба сезона была полидоминантной - преобладали сапрофаги: весной - дождевые черви (40%) и энхитреиды (9%), осенью - личинки двукрылых и щелкунов (21-26%) и энхитреиды (7%). Трофическая структура значительно изменялась по сезонам из-за качественных и количественных перестроек в трофической группе насекомых-фитофагов. В комплексах сапро- и зоофагов происходило сезонное перераспределение доминирования по численности и биомассе между одними и теми же таксонами. Весной почти половину общей численности составляли фитофаги (с долей щитовок 90%) и сапрофаги (с долей энхитреид 50% и двукрылых 30%). По биомассе в оба сезона доминировали сапрофаги, но с меньшей долей, чем в субальпийском поясе: 62-66% против 71-72(табл. 5).
5.3. Пояс полярной пустыни (горное плато с фрагментарной травянисто-мохово-лишайниковой растительностью)
На предельных для Хибинского горного массива высотах более 1 тыс. м над ур. м. выявлено 14 таксонов беспозвоночных. В оба сезона под подушками растительности встречались сапрофильные энхитреиды и личинки двукрылых, бриофаги пилюльщики, пауки, многоножки, жужелицы, стафилиниды и долгоносики (всего 8 таксонов). В высотном ряду г. Ловчорр число таксонов было наименьшим из-за отсутствия ряда сапро- и фитофагов. Здесь не выявлены дождевые черви и моллюски, а из насекомых - щелкуны, клопы, чешуекрылые и трипсы. Обнаружено 4 вида стафилинид и 2 голарктических вида жужелиц: бореальный миксофитофаг Amara brunnea и тундровый зоофагPterostichus brevicornis. Европейско-средиземноморский тундрово-лесной вид Mycetoporus longulus (Mannerheim, 1831), выявленный в пределах Хибин только в полярной пустыне, является новыми для Мурманской обл., а вид Euaesthetus sp. представляет новое для Хибин п/сем. Euaesthetinae.
Численность (~200 экз./м2) и биомасса (~400 мг/м2) были наименьшими в высотном ряду г. Ловчорр. В оба сезона супердоминировали личинки двукрылых, как и в ниже лежащих поясах. Их численность достигала 60-140 экз./м2, а обилие было максимальным по склону горы (47-63%).
На фоне низкой плотности большинства таксонов возросла значимость хищных многоножек вида Monotarsobius curtipes (C.Koch, 1847) до 17% при численности 22-37 экз./м2 в сравнении с 3-5 экз./м2 (или менее 2%) в подстилках субальпийского пояса и 10-37 экз./м2 (или 2-5%) в подстилках альпийского пояса г. Ловчорр. Увеличение численности этого вида в горно-тундровых сообществах по сравнению с горно-лесными отмечено и на г. Вудъяврчорр.
По биомассе в оба сезона преобладали хищные многоножки (40%) и детритоядные личинки пилюльщиков (27), а весной еще и личинки двукрылых и пауки (13-14). Осенью преобладали пилюльщики (61) и многоножки (21). Трофическая структура отличалась резко обедненным составом сапро- и фитофагов, низкими значениями численности и биомассы всех трофических групп и сезонной сменой доминирования сапро- и зоофагов. (табл. 5). Комплекс зоофагов в разные сезоны был наиболее стабильным по численности и биомассе и включал те же группы, что и в расположенных ниже по склону поясах.
ГЛАВА 6. ОБЩИЕ ЗАКОНОМЕРНОСТИ ОРГАНИЗАЦИИ СООБЩЕСТВ БЕСПОЗВОНОЧНЫХ В ПОЧВАХ ХИБИНСКИХ ГОР
Методами почвенных раскопок и ловушек выявлено 29 таксонов: от 8 до 24 в разных биоценозах при наибольшем разнообразии в лесных поясах всех гор. Большинство таксонов одинаково хорошо учитывались обоими методами. Только ловушками учтены скорпионовые мухи (Mecopterа). Только при раскопках в подстилках березняков на южных склонах гор выявлены геобионтные многоножки-землянки (Geophilidae), редких в области, и, вероятно, не обитающих севернее границы леса. Общими для трех гор были 16 таксонов.
В большинстве биоценозов независимо от сезона преобладали беспозвоночные с короткими циклами развития в почвенной среде - личинки двукрылых и энхитреиды. Доля двукрылых варьировала от 18 до 77%, доля энхитреид от 5 до 70%, достигая максимальных значений в подстилках субальпийского и альпийского поясов. В большинстве биоценозов независимо от сезона преобладали беспозвоночные с короткими циклами развития в почвенной среде - личинки двукрылых и энхитреиды. Доля двукрылых варьировала от 18 до 77%, доля энхитреид от 5 до 70%, достигая максимальных значений в подстилках субальпийского и альпийского поясов. На открытых пространствах горной тундры возрастали численность и обилие хищников - пауков и многоножек-костянок (до 10-17%). Весной в горных тундрах массово размножались почвообитающие партеногенетические щитовки, для которых нами отмечено как вынашивание яиц, так и не характерное для этих насекомых живорождение.
Хибинские тундры отличались от зональной кольской тундры высокой численностью и разнообразным составом почвенной фауны (13-17 таксонов против 5-10), что можно объяснить более мягкими климатическими условиями в связи с расположением Хибин в подзоне северной тайги, а самих тундр - на небольших высотах. Во всех тундрах доминируют двукрылые, жесткокрылые и энхитреиды, но плотность этих групп и общая численность беспозвоночных в горных тундрах сопоставима с зональными сосняками (табл. 7).
Табл. 7. Показатели почвенной фауны в горных и зональных биоценозах Мурманской области
Биоценозы |
Горные |
Зональные |
||||
Ельники n = 2 |
Березняки n = 5 |
Тундры n = 6 |
Полярная пустыня |
Ельники n = 5 |
Сосняки n = 5 |
|
Числотаксонов |
17-20 |
16-20 |
13-17 |
13 |
15-23 |
14-25 |
Численность, экз./м2 |
430-560 |
370-600 |
180-800 |
180 |
560-710 |
250-550 |
Биомасса, г/м2 |
2.5-4.0 |
0.9-11.3 |
0.2-4.8 |
0.4 |
0.8-1.6 |
0.2-10.0 |
Почвенная фауна лесных поясов Хибин (горно-таежные ельники и горно-лесотундровые березняки), напротив, представляет обедненный вариант педофауны северной тайги. По сравнению с зональными сосняками и ельниками в подстилках горно-лесных биоценозов отсутствует ряд таксонов, плотность доминирующих отрядов (двукрылые, жесткокрылые, пауки) ниже, а олигохет (люмбрициды, энхитреиды) - выше (табл. 8). Высокая доля гумификаторов - люмбрицид и энхитреид в составе сапрофильного комплекса отличает высоко гумусированные и обогащенные биогенными элементами (Ca, K, Si, Mg, P, S) горные почвы Хибин от северотаежных подзолов и свидетельствуют о большей функциональной активности сапроблока в горных почвах по сравнению с зональной тайгой Мурманской области.
Табл. 8. Соотношение основных таксонов беспозвоночных в горных и зональных биоценозах Мурманской области (обилие, %)
Беспозвоночные |
Горные биоценозы |
Зональные |
|||
Ельники |
Березняки |
Тундры |
Полярная пустыня |
Ельники, сосняки |
|
Насекомые |
48-65 |
11-86 |
22-85 |
73 |
85-89 |
Черви |
12-47 |
8-75 |
3-71 |
9 |
7-9 |
Паукообразные |
2-17 |
2-10 |
0-12 |
1 |
3-5 |
Многоножки |
1-4 |
1-7 |
0-7 |
17 |
1-2 |
Моллюски |
2 |
0-2 |
0-2 |
0 |
1 |
Итого, % |
100 |
100 |
100 |
100 |
100 |
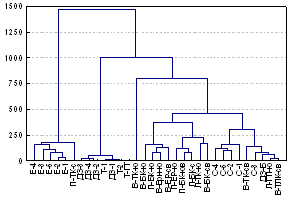
Различия в составе и численности таксонов, преобладающих в горных и зональных почвах, отразились в большем фаунистическом сходстве между биоценозами Хибин, чем между горными и зональными биоценозами (рис. 2).
На численность беспозвоночных в горных биоценозах достоверное влияние оказывают следующие факторы (в порядке убывания): горно-растительный пояс (F=8.1, p=0.0005) при min значениях средней численности в арктической пустыне и max в альпийском поясе > гора (F=5.2, p=0.02), min на г. Ловчорр, max на г. Поачвумчорр > высота над уровнем моря (F=3.7, p=0.0003), min на высоте 1093 м, max на высотах 590-600 м. На уровень биомассы влияет фактор гора (F=10.3, p=0.0001), min на г. Ловчорр, max на г. Поачвумчорр > лэкспозиция склона (F=4.2, p=0.01), min на северных склонах, max на южных.
Ward`s method, Euclidean distances, 32 variants |
Рис. 2. Дендрограмма фаунистического сходства горных и зональных биоценозов по разнообразию таксонов и численности почвенной фауны. Зональная тундра: ДЗ1-3, ДЗ-Б - окрестности пос. Д. Зеленцы, Т1-3, Т-ГТ - окрестности пос. Териберка; обозначение остальных биоценозов - как в табл. 1 и на рис. 1. |
ГЛАВА 7. ВИДОВОЕ РАЗНООБРАЗИЕ БЕСПОЗВОНОЧНЫХ В ПОЧВАХ ХИБИНСКИХ ГОР
Оценено разнообразие ключевых сапрофагов - дождевых червей и жуков трех семейств (стафилиниды, жужелицы, щелкуны), наиболее массовых в экосистемах гор.
7.1. Жесткокрылые (Coleoptera: Staphylinidae, Carabidae, Elateridae)
7.1.1. Общая характеристика разнообразия доминирующих семейств
Методами почвенных проб и ловушек выявлено 79 видов: 47 видов стафилинид из 27 родов, 7 п/сем.; 22 вида жужелиц из 13 родов, 4 п/сем.; 10 видов щелкунов из 10 родов, 4 п/сем. Впервые для Хибин указываются 42 вида стафилинид, 11 видов жужелиц и 6 видов щелкунов, впервые для Мурманской области - 19, 4 и 1 вид из этих семейств.
Наименее разнообразна фауна жуков г. Вудъяврчорр (30 видов: стафилинид 18, жужелиц 8, щелкунов 4 вида) из-за расположения большинства биоценозов на северо-восточном склоне. На г. Поачвумчорр и Ловчорр выявлено 54 и 46 видов жуков. Стафилиниды были наиболее разнообразны в лесных биоценозах г. Поачвумчорр: 21-24 вида из 13-15 родов (всего 33 вида, 18 родов). Жужелицы - на северных склонах: в субальпийском поясе г. Ловчорр (14 видов из 10 родов) и в тундровых биоценозах г. Поачвумчорр (10 видов из 7 родов), как и щелкуны (по 6 видов из 6 родов в каждом из этих биоценозов). Во всех горных экосистемах видовой спектр стафилинид был шире по сравнению с жуками других семейств. Общими для 3 гор были лишь 13 из 79 видов жуков (или 17%): по 5 видов стафилинид и жужелиц и 3 вида щелкунов.
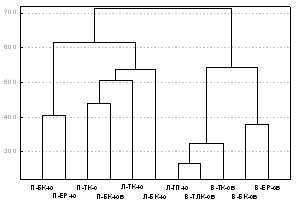
Фаунистическое своеобразие жесткокрылых и, следовательно, различие экологических условий на разных горах подтверждено низкими значениями коэфф. Съеренсена (<22%) и результатами кластеризации данных (рис. 3). В один кластер (справа) выделились биоценозы северо-восточного склона г. Вудъяврчорр, в другой кластер - биоценозы гор Ловчорр и Поачвумчорр, расположенные в основном на склонах, ориентированных к югу. В пределах этих кластеров обособились горно-лесные биоценозы соседних поясов каждой горы и биоценозы субальпийского и альпийского поясов, что свидетельствует о взаимном проникновении таежных и тундровых видов жуков в экосистемы этих поясов.
|
Рис. 3. Убывание сходства видового состава жесткокрылых (суммарно по трем семействам) в ряду: соседние горно-растительные пояса на склоне одной горы (коэф. Съеренсена 38-49%) > одноименные горно-растительные пояса на разных горах (7-36%) > одноименные горно-растительные пояса на разных склонах одной горы (13-15%). |
Для 14 биоценозов Хибин ни одного общего вида жуков из трех семейств не выявлено. Из 79 видов 46 (или 58%) встречались локально - лишь в 1-2 горных биоценозах, в т.ч. 29 видов (или 60%) стафилинид, 12 видов (55%) жужелиц, 5 видов (50%) щелкунов. Около половины всех жуков (31 вид или 39%), в т.ч. 16 видов стафилинид, 10 видов жужелиц и 5 видов щелкунов были приурочены к определенному растительному поясу. Наибольшее число видов каждого семейства (стафилинид 10, жужелиц 5, щелкунов 4) как лесной, так и тундровой преференции, населяли переходный субальпийский пояс, что можно рассматривать как проявление экотонного эффекта.
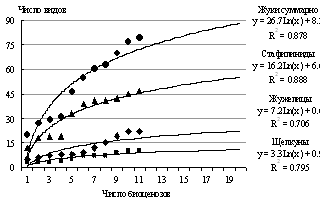
С учетом сильной расчлененности горного ландшафта и гетерогенности местообитаний, определяющих локальную встречаемость видов, можно прогнозировать обнаружение новых видов жуков. Кривые зависимости насыщения видовых списков от числа исследованных биоценозов [Коросов, 2007] показывают, что обследование 17 биоценозов (т.е. 5 гор) позволит учесть 99% видового разнообразия 3 семейств, что составляет около 90 видов (рис. 4).
|
Число |
|||
гор |
биоценозов |
видов |
% |
|
3 |
11 |
79 |
94 |
|
4 |
14 |
84 |
98 |
|
5 |
17 |
88 |
99 |
|
Рис. 4. Увеличение числа видов жуков в горах с увеличением числа обследованных биоценозов. |
||||
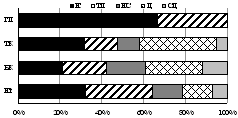
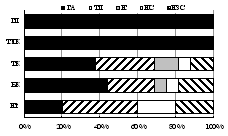
7.1.2. Семейство Стафилиниды (Staphylinidae)
В почвах трех гор выявлено 47 видов стафилинид из 27 родов, 7 п/сем. Наиболее разнообразны п/сем. Aleocharinae (19 видов) и Tachyporinae (10), адаптированные к обитанию в почвах северной тайги, тундры и горно-альпийских экосистемах [Колесникова, Ужакина, 2005]. По числу видов преобладали лишь 4 рода: Atheta (7 видов), Quedius (4),Oxypodaи Tachinus (по 3). Большинство родов (23 рода или 85%) включали по 1-2 вида, что отражает миграционный характер формирования горной фауны. По численности доминировали виды п/сем. Xantholininae и Aleocharinae: Xantholinuslinearis, Othiuslapidicola, Athetabrunneipennis, Drusillacanaliculata, Deviaprospera. Доминирование отдельных видов в биоценозах горно-таежного и альпийского поясов не превысила 30% при разнообразии 12-20 видов. В экосистемах субальпийского пояса на фоне высокого числа видов (до 24) обилие эвритопного D. canaliculata достигало 40%.
Установлено влияние поясности на видовой состав, зоогеографическую и экологическую структуру комплексов стафилинид. Лесные по биотопическому преферендуму Deliphrumtectum, Atheta myrmecobia, Lioglutagranigera, Lioglutamicans, Oxypodaskalitzkyi иO. sororвстречены лишь в таежном поясе гор. Еще 11 лесных, луговых и эвритопных видов обитали в горно-лесных поясах, не проникая в тундры. Только в субальпийских березняках выявлены 10 видов стафилинид лесной и тундровой преференции; только в альпийском поясе - лесные и тундровые Haploglossavillosula, Mniusaincrassata, Acidotaquadrata, Anthophagusalpinus. В каменистом субстрате суровой арктической пустыни г. Ловчорр на высоте 1100 м обнаружены 4 вида, 3 из которых Atheta fungi, A. brunneipennis и Euaesthetus sp. являются новыми для Хибин, а вид Mycetoporus longulus, известный для высокогорных экосистем мира - новым для Мурманской области.
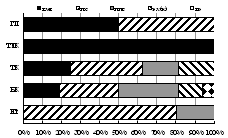
В целом сообщества этих жуков наиболее разнообразны в лесных поясах Хибин (27-34 вида против 4-18 в тундровых) и наиболее сбалансированы по зоогеографической и экологической структуре в субальпийском поясе (рис. 5).
Для Хибин было известно 15 видов стафилинид [Россолимо, 1994], из них нами не обнаружены 9 видов. С учетом всех данных в горных почвах обитает не менее 56 видов из 28 родов, 7 п/сем.. По числу видов преобладают рода Atheta (7 видов), Quedius,Oxypodaи Tachinus (по 3-4). Фауна стафилинид Мурманской обл. насчитывает 187 видов из 64 родов и 12 подсемейств при наибольшем разнообразии родов Stenus (23 вида), Atheta (15), Quedius, Philonthus (12-13) и Lathrobium (9).
I |
II |
III |
Рис. 5. Зоогеографическая (I), трофическая (II) и морфо-экологическая (III) структура комплексов стафилинид в высотных поясах Хибин. I: - Ц циркумареал, СЦ - субциркумареал, ТП - транспалеарктический, ЕС - европейско-сибирский, Е - европейский. II: с - сапрофаг, з - зоофаг, зм - зоомицетофаг, фд - фитодетритикол. III: сх - стратохортобионт, сп - стратобионт подстилочный, спп - стратобионт поверхностно-подстилочный, сппч - стратобионт подстилочно-почвенный. |
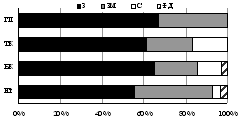
7.1.3. Семейство Жужелицы (Carabidae)
Выявлено 22 вида жужелиц из 13 родов 4 п/сем. Новыми для Хибин и региона в целом оказались 4 вида: Общими для трех гор были лишь 5 видов: Amara brunnea, Calathus melanocephalus, C. micropterus, Pterostichus brevicornis, Patrobus assimilis. Они встречались как в лесных, так и тундровых поясах Хибин. Максимальное число видов (10-14) характерно для альпийского пояса г. Поачвумчорр и субальпийского пояса г. Ловчорр. Наименьшее разнообразие (1-3 вида) отмечено в верхнем поясе арктической тундры г. Вудъяврчорр, полярной пустыне г. Ловчорр и лесных поясах южного склона г. Поачвумчорр. Наиболее высотное распространение в Хибинах (1093 м над ур. м.) установлено для жужелиц A. brunnea и P. brevicornis. Оба вида имеют обширные голарктические ареалы, а в Хибинах обитают в субальпийских и альпийских поясах всех гор.
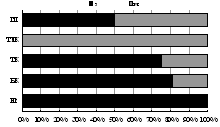
Изменению числа видов жужелиц в высотно-поясном градиенте (5>16>15>2) соответствует смена зоогеографической и экологической структуры их сообществ (рис. 6). Большинство видов, обитающих в лесных и тундровых поясах Хибин, являются зоофагами по типу питания и принадлежат к жизненной форме подстилочных или поверхностно-подстилочных стратобионтов. С высотой наблюдается смена преобладающей жизненной формы в ряду: подстилочные стратобионты > поверхностно-подстилочные стратобионты > геохортобионты. Хищные жужелицы вытесняются видами-фитофагами. Из 7 видов, встреченных в наиболее высотных поясах, 5 видов жужелиц в стадии имаго являются миксофитофагами, способными обитать в ярусе напочвенной растительности. Наиболее разнообразные сообщества жужелиц при сбалансированном соотношении зоогеографических и экологических групп характерны для субальпийских и альпийских поясов Хибин.
I |
II |
III |
Рис. 6. Зоогеографическая (I), трофическая (II) и морфо-экологическая (III) структура комплексов жужелиц в высотных поясах Хибин. I: ГА - голарктический, ТП - транспалеарктический, Е - европейский, ЕС - европейско-сибирский, ЕЗС - Европейско-западносибирский; II: пс - подстилочный стратобионт, ппс - поверхностно-подстилочный стратобионт, эх (к) - эпигеобионт ходящий (крупный), гр - геобионт роющий, с - стратобионт, гх-г - геохортобионт гарпалоидный. III: з - зоофаг, м - миксофитофаг. |
В исследованных биоценозах трех гор нами не были обнаружены 6 видов жужелиц из 8 родов, указанные для Хибин в предыдущих работах [Россолимо, 1989, 1994; Эколого-экономическое обоснованиеЕ, 1999]; с их учетом карабидофауна Хибин насчитывает не менее 30 видов из 17 родов, а Мурманской обл. - 69 видов жужелиц из 26 родов относящихся к 8 п/сем. при наибольшем разнообразии родов Amara (17 видов), Bembidion и Agonum (по 7). В Хибинах по числу видов преобладают мелкоразмерные подстилочные хищники рода Notiophilus и травоядные жужелицы мелких и средних размеров из рода Amara (по 4 вида).
В исследованных биоценозах трех гор нами не были обнаружены 6 видов жужелиц из 8 родов, указанные для Хибин в предыдущих работах [Россолимо, 1989, 1994; Эколого-экономическое обоснование Е, 1999]: Amara (=Paracelia) quenseli (Schonherr, 1806), Bembidion bipunctatum (Linnaeus, 1761), B. yukonum (Fall, 1926), Bradycellus caucasicus (Chaudoir, 1846), Cymindis vaporariorum (L., 1758), Elaphrus riparius (L., 1758), Loricera pilicornis (Fabricius, 1755). С учетом этих видов карабидофауна Хибин насчитывает не менее 30 видов из 17 родов, а Мурманской области в целом - 69 видов жужелиц из 26 родов относящихся к 8 подсемействам при наибольшем видовом разнообразии родов Amara (17 видов), Bembidion и Agonum (по 7). В Хибинах по числу видов наиболее разнообразны мелкоразмерные подстилочные хищники рода Notiophilus и травоядные жужелицы мелких и средних размеров из рода Amara (по 4 вида).
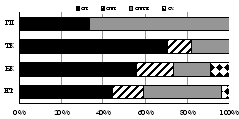
7.1.4. Семейство Щелкуны (Elateridae)
В биоценозах Хибин идентифицировано 10 видов щелкунов из 10 родов, 7 из них относятся к всесветно распространенному п/сем. Athoinae, разнообразному в горных экосистемах. Еще 3 вида родов Oedostethus и Limonius, предположительно виды O. nubilus, O. curticornis и L. parallelus, отловленные почвенными ловушками по 1 экз. в субальпийских поясах гор Ловчорр и Поачвумчорр, требуют уточнения. Щелкуны, ранее известные для Мурманской обл., но впервые выявленные в Хибинах, также отловлены в субальпийских поясах - березняке травянистом на южном склоне г. Поачвумчорр (лесные Mosotalesus impressus иDalopius marginatus) и редкостойном березняке на северном склоне г. Ловчорр (борео-монтанный Orithales serraticornis и аркто-борео-монтанный Hypnoidus rivularius). В этом же березняке обнаружены мелкие гигрофильные: щелкун северный Ascoliocerus (Berninelsonius) hyperboreus, широко распространенный в зональных и горных тундрах Евразии, но новый для региональной фауны, и щелкун прибрежный Negastrius pulchellus, известный в Хибинах из работ В.Ю. Фридолина [1936], но не найденный в других районах Мурманской обл.
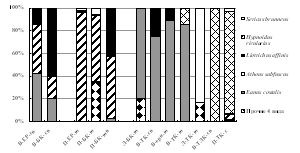
Аркто-альпийские и аркто-борео-монтанные N. pulchellus, A. hyperboreus и H. rivularius, способные выдержать суровые условия перигляциальной зоны, считаются пионерными видами щелкунов на северных территориях после отступления ледников [Медведев, 2005]. Бореальные и температные виды, населяющие горно-лесные пояса Хибин и составляющие основу региональной фауны элатерид, распространялись на север по мере потепления климата и появления лесов по интразональным биотопам из Европы и Сибири. Таковы борео-монтанные трансевразиатские Liotrichus affinis, Orithales serraticornis, Mosotalesus impressus, температный трансевразиатский Sericus brunneus, виды европейско-сибирской группы - евро-байкальский D. marginatus и евро-обский A. subfuscus. Наиболее широко в Хибинах распространены бореальные и температные виды E. costalis,A. subfuscus, L. affinis и S. brunneus, фоновые в подстилках зональных сосняков и ельников (рис. 7). Общими для трех гор оказались лишь 3 вида щелкунов из 10: H. rivularius, E. costalis,L. affinis). Преобладание E. costalis в тундровых и лесотундровых биоценозах разных гор соответствует его характеристике как наиболее холодоустойчивого вида щелкунов, личинки которого имеют крайне низкие температуры переохлаждения: до -12 Е-16 С [Россолимо, 1994].
|
Рис. 7. Структура элатеридокомплексов в экосистемах Хибин. |
Проволочники большинства видов принадлежат к подстилочной экологической группе, их личиночное развитие связано с лесными местообитаниями. Это отражает таежный облик хибинской фауны щелкунов. По типу питания преобладают виды, которые сочетают сапро-, некрофагию и необлигатное хищничество и не повреждают живые растения. Имаго щелкунов, обитающих в Хибинах, ведут открытый образ жизни и активны в дневное время суток. Виды с сумеречно-ночной и смешанной активностью (самцы активны в течение суток, самки - только в сумерки или ночью) единичны в зональных биоценозах (роды Elater, Agriotes) и не встречены в горных.
С учетом наших данных элатеридофауна Хибин насчитывает не менее 10 видов из 10 родов, а Мурманской обл. в целом - 40 видов из 22 родов относящихся к 4 п/сем. при наибольшем видовом разнообразии родов Selatosomus и Ampedus (по 5 видов).
7.2. Дождевые черви (Oligochaeta: Lumbricidae)
В биоценозах Хибин известны 4 вида дождевых червей (D. octaedra, D. rubudus, A. caliginosus и L. rubellus): от 3-4 видов с численностью до 130 экз./м2.в межгорных березняках до одного (D. octaedra) на вершинах гор [Рыбалов, 2006]. Нами выявлено 6 видов в основном почвенно-подстилочной жизненной формы: в горных ельниках 4, в лесотундровых березняках 5, в альпийских тундрах 6 видов (табл. 9).
Спектр видов и жизненных форм червей варьировал как по горам и склонам, так и по растительным поясам при наибольшем разнообразии в субальпийских березняках на южных склонах. Лиственным лесам с большими запасами подстилки, ее меньшей кислотностью и повышенным по сравнению с горными ельниками содержанием минеральных элементов, углерода и азота [Морозова и др., 1976; Почвообразование Е, 1979] соответствовали высокие показатели численности и биомассы червей: до 55 экз./м2 и 8 г/м2 на г. Поачвумчорр, до 70 экз./м2 и 12 г/м2 на г. Вудъяврчорр. Обитание крупного теплолюбивого вида E. fetida в субальпийских березняках на склонах южной и юго-восточной экспозиции свидетельствует об интразональном характере этих биоценозов и полугидроморфном режиме горных почв.
Горно-лесные пояса Хибин (таежный и лесотундровый) характеризовались разнообразием морфо-экологических типов червей, которые образуют сукцессионный ряд: подстилочные > подстилочно-почвенные > почвенные. Он соответствует стадийности процесса деструкции растительного опада [Hendrix & Bohlen, 2002; Eisenhauer et al., 2007] и является показателем более глубокой степени биотрансформации органического вещества в почвенном ярусе горно-лесных биоценозов по сравнению с зональными подзолами.
Табл. 9. Обилие (%) дождевых червей в почвах высотных поясов Хибин
Гора |
Вудъяврчорр |
Поачвумчорр |
овчорр |
|||||||||
Пояс |
ЕР |
БК |
ТК |
ТЛК |
ЕР |
БК |
БК |
ТК |
БК |
ТК |
ГП |
|
Экспозиция |
св |
ю |
юв |
с |
с |
ю |
|
|||||
Питающиеся растительным опадом на поверхности почвы |
||||||||||||
есные подстилочные |
||||||||||||
Dendrobaena octaedra (Sav.) |
Ц |
35 |
20 |
Ц |
16 |
8 |
10 |
100 |
100 |
Ц |
Ц |
|
Почвенно-подстилочные |
||||||||||||
Dendrodrilus rubidus (Sav.) |
Ц |
8 |
20 |
Ц |
84 |
16 |
80 |
Ц |
Ц |
Ц |
Ц |
|
Lumbricus rubellus (Hoff.) |
50 |
57 |
20 |
Ц |
Ц |
20 |
8 |
Ц |
Ц |
Ц |
Ц |
|
Eisenia fetida (Sav.) |
Ц |
Ц |
Ц |
Ц |
Ц |
20 |
2 |
Ц |
Ц |
Ц |
Ц |
|
Eisenia nordenskioldi(Eisen) |
Ц |
Ц |
20? |
Ц |
Ц |
Ц |
Ц |
Ц |
Ц |
Ц |
Ц |
|
Питающиеся почвенным перегноем |
||||||||||||
Собственно-почвенные среднеярусные |
||||||||||||
Aporrectodea c. caliginosa (Sav.) |
50 |
Ц |
20 |
Ц |
Ц |
36 |
Ц |
Ц |
Ц |
Ц |
Ц |
|
Всего |
видов |
2 |
3 |
4+1? |
0 |
2 |
5 |
4 |
1 |
1 |
0 |
0 |
экземпляров |
2 |
40 |
5 |
0 |
58 |
30 |
186 |
3 |
3 |
0 |
0 |
|
На северо-восточном склоне г. Вудъяврчорр максимальное в высотно-поясном ряду разнообразие люмбрицид (5 видов) выявлено весной в тундре кустарничковой на высоте 370 м, при этом все виды были представлены единичными особями. Азиатский по происхождению вид E. nordenekioldi найден только в этом биоценозе и указывается для Мурманской обл. впервые.
ВЫВОДЫ
1. Почвенная фауна Хибин представлена не менее чем 28 таксонами. В ельниках горной тайге выявлено от 20 до 24 таксонов, в лесотундровом поясе 9-24, в тундрах 8-22. По численности во всех поясах преобладают насекомые (11-85%) и черви (8-71%), главным образом, двукрылые (18-77%) и энхитреиды (до 70%) с короткими циклами развития в почве. Доля паукообразных и многоножек не превышает 17%, моллюсков - 2%. С высотой снижается доля тонкопокровных червей и моллюсков и возрастает обилие членистоногих (многоножек и паукообразных).
2. Почвенная фауна горно-лесных поясов сохраняет ядро доминантов, характерных для подзолов зональной северной тайги (двукрылые, жесткокрылые, пауки, многоножки, энхитреиды, люмбрициды), но обеднена насекомыми разных трофических уровней. Горные тундры, напротив, отличаются от зональной тундры высокой численностью и разнообразием почвенной фауны, сопоставимым с зональной тайгой. Это объясняется мягкими климатическими условиями альпийского пояса в связи с положением Хибин в подзоне северной тайги, а самих тундр - на небольших высотах. О мягкости горного климата свидетельствует таежный облик фауны жесткокрылых и проникновение лесных видов жуков в альпийский пояс.
3. В высоко гумусированных и обогащенных биогенными элементами почвах Хибин хорошо развит комплекс сапрофагов с преобладанием ключевых гумификаторов - дождевых червей и энхитреид, что свидетельствуют о большей функциональной активности сапроблока в горных экосистемах по сравнению с зональной тайгой. Показателем более глубокой степени биотрансформации органического вещества в почвенном ярусе горно-лесных биоценозов является широкий спектр видов и морфо-экологических типов червей, которые образуют сукцессионный ряд: подстилочные > подстилочно-почвенные > почвенные, соответствующий стадийности процесса деструкции опада. В субальпийских березняках южных склонов высокие показатели численности (до 500 экз./м2), биомассы (до 12 г/м2) и разнообразия сапрофагов, отличающие структуру высокопродуктивных сообществ, свидетельствуют об интразональном характере этих биоценозов.
4. Фактор поясности в большей степени влияет на сапро- и фитофагов, в меньшей - на хищников. Среди зоофагов перераспределяется доминирование по численности и биомассе между таксонами при сохранении их состава от 5-7 таксонов в горно-лесных поясах до 6-7 в тундрах. Сапрофильный комплекс в горных тундрах обедняется крупноразмерными дождевыми червями и моллюсками (3-4 таксона против 6-7 в горно-лесных поясах). В комплексе фитофагов с высотой сужается спектр таксонов, насекомые-обитатели напочвенной растительности сменяются почвенно-подстилочными видами щитовок и жуками - пилюльщиками, листоедами, долгоносиками. Для ряда групп (жесткокрылые, многоножки, дождевые черви) высотно-поясное распределение сходно с широтно-зональным распространением в пределах региона. Для массовых групп жуков (стафилинид, жужелиц, щелкунов) подтверждена смена таежных видов тундровыми при переходе от таежного пояса к субальпийскому и, далее, к альпийскому. Во всех поясах по составу и численности преобладают виды с лесной преференцией.
5. Влияние экспозиции склона проявляется в более высокой функциональной активности сапроблока и достоверно высокой численности сапрофагов (люмбрицид, энхитреид, моллюсков, элатерид) в почвах южных склонов по сравнению с зональной северной тайгой. Почвенная фауна склонов северной и северо-восточной экспозиции по составу таксонов и численности ближе к таковой в зональных подзолах.
6. Период активной трофической деятельности сапрофагов в горных почвах ограничен 3-4 месяцами. Наиболее уязвимы к воздействию природно-климатических условий Хибин крупноразмерные тонкопокровные дождевые черви, моллюски и аэробионтные насекомые, личинки которых развиваются в почве. Экстремальными горными факторами, влияющими на численность, разнообразие и сезонную активность фауны являются: крутизна склонов, нестабильный гидротермический режим, значительная скорость ветра, короткий период с положительными температурами воздуха.
7. В горных биоценозах выявлено 79 видов жесткокрылых: 47 видов стафилинид из 27 родов, 7 п/сем.; 22 вида жужелиц из 18 родов, 4 7 п/сем.; 10 видов щелкунов из 10 родов, 7 п/сем. Впервые для Хибин указаны 42, 11 и 6 видов из этих семейств, впервые для Мурманской области - 19, 4 и 1 вид. Региональный список стафилинид расширен до 187 видов из 64 родов, жужелиц - до 69 из 26 родов, щелкунов - до 40 видов из 22 родов. С учетом литературных данных хибинская фауна стафилинид, жужелиц и щелкунов теперь насчитывает 56, 30 и 10 видов соответственно. Типичными обитателями Хибин можно считать лишь 12 видов, наиболее широко распространенных в горных биоценозах: стафилинид Mycetoporus lepidus, M. monticola, Atheta brunneipennis, A. graminicola, Oxypoda annularis, жужелиц Patrobus assimilis, Calathus micropterus, Amara brunnea, щелкуны Eanus costalis, Liotrichus affinis, Athous subfuscus, Hypnoidus rivularius.
Список публикаций по теме исследования
Публикации в изданиях, рекомендованных ВАК РФ
Зенкова И.В., Пожарская В.В., Похилько А.А. Материалы к почвенной фауне Хибинского горного массива на примере горы Вудъяврчорр // Вестник МГТУ, 2009. Т. 12. № 3. С. 516-524.
Зенкова И.В., Пожарская В.В., Похилько А.А. Высотное распределение почвенной фауны Хибин // Почвоведение, 2011. №9. С. 1083-1093.
Зенкова И.В., Пожарская В.В., Филиппов Б.Ю., Колесникова А.А., Середюк С.Д. Высотная поясность и комплексы жесткокрылых в почвах Хибинского горного массива // Труды Карельского НЦ РАН, 2011. №2. С. 107-118.
Публикации в тематических сборниках и трудах конференций
Зенкова И.В., Пожарская В.В. Беспозвоночные животные в почвах горно-растительных поясов Хибин (на примере г. Вудъяврчорр) // Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт / Матер. междун. конф. - Мурманск: МГПУ, 2009. С.101-105.
Пожарская В.В., Зенкова И.В. Комплексы беспозвоночных в подстилках равнинных и горного ельников Мурманской области // Продуктивность и устойчивость лесных почв / Матер. междун. конф. по лесному почвоведению. - Петрозаводск, 2009. С. 218-220.
Пожарская В.В., Зенкова И.В. Комплексы жуков-щелкунов (Сем. Elateridae) в равнинных и горных почвах северотаежной подзоны Мурманской области // Энтомологические исследования в Северной Азии / Матер. VIII межрегион. совещ. энтомологов Сибири и Д. Востока. - Новосибирск, 2010. С. 169-171.
Пожарская В.В., Зенкова И.В. Жесткокрылые (Coleoptera: Stafilinidae, Carabidae, Elateridae) в почвах Хибин // Экологические функции лесных почв в естественных и нарушенных ландшафтах / Матер. IV Всерос. научн. конф. по лесному почвоведению. - Апатиты: Кольский НЦ РАН, 2011. Ч. 1. С. 44-48.
Зенкова И.В., Пожарская В.В. Разнообразие беспозвоночных животных в почвах тундровой и лесной зоны Мурманской области / Матер. всерос. конф. к 100-летию И.П. Щербакова. - Якутск, 2011. С. 104-110.
Пожарская В.В., Зенкова И.В. Почвенная мезофауна Хибинского горного массива (Мурманская область) // Проблемы почвенной зоологии / Матер. XVI всерос. совещ. по почвенной зоологии. - Москва, 2011. С. 90-92.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]