
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
ПРИБРЕЖНОЕ СООБЩЕСТВО ОБРАСТАНИЯ В РАЗНЫХ ЭКОЛОГИЧЕСКИХ ЗОНАХ КАСПИЙСКОГО МОРЯ
Автореферат кандидатской диссертации
а аНа правах рукописи
ИМАШОВА САНИЯТ НИГМАТУЛАЕВНА
Прибрежное сообщество обрастания в разных экологических зонах КАСПИЙСКОГО МОРЯ
03.02.08 - экология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
![]() МАХАЧКАЛА - 2012
МАХАЧКАЛА - 2012
Работа выполнена в лаборатории гидробиологии и химической экологии моря Прикаспийского института биологических ресурсов Дагестанского научного центра Российской академии наук.
Научный руководитель: а доктор биологических наук,
Зайко Владимир Алексеевич
Официальные оппоненты: доктор биологических наук,
Заслуженный рыбовод Российской Федерации,
профессор кафедры Ихтиология ДГУ,
Магомаев Феликс Магомедович
доктор биологических наук, профессор,
Почетный работник рыбного хозяйства
РФ Председатель Астраханского отделения
Всероссийского общества охраны природы
Сокольский Аркадий Федорович
Ведущая организация: Астраханский государственный технический университет
Защита диссертации состоится 10 апреля 2012г. в 14.00ч. на заседании Совета по защите докторских и кандидатских диссертаций Д212.053.03 при Дагестанском государственном университете по адресу: 367025, г.Махачкала, ул. Дахадаева, 21.
С диссертацией можно ознакомиться в библиотеке ФГБОУ ВПО Дагестанский государственный университет
Автореферат разосланаа аа_______________________
Ваш отзыв, заверенный печатью, просим направлять по адресу:
367025, г.Махачкала, ул. Дахадаева, 21.
факс: 8(8722) 67-46-51
e-mail: есоdag@rambler.ru
Ученый секретарь
Диссертационного Совета,а
к.г.н., доцент Ахмедова Г.А.
ВВЕДЕНИЕ
Актуальность. Изучение процессов формирования и функционирования морских сообществ необходимо, поскольку такие исследования позволяют получить информацию о направлениях и темпах развития экосистем и их деградации в естественных условиях и при антропогенных стрессовых воздействиях. Отсутствие или фрагментарность данных о скоростях и направлениях сукцессии многообразных прибрежных сообществ обрастания, об общих тенденциях и закономерностях изменчивости и устойчивости различных морских экосистем значительно сдерживают разработку методологии рационального использования ресурсов Мирового океана. Особое значение эти исследования приобретают на Каспии в связи с проводящимися и планируемыми работами по добыче углеводородов.
Анализ сложившихся представлений о сообществах аобрастания показал, что мы имеем дело с крайне разнородными ценозами (по составу и структуре, сезонной и многолетней динамике). Существует необходимость в решении задачи их упорядоченного описания.
Цель исследования: выявить особенности существования прибрежного сообщества обрастания в разных экологических зонах моря в связи с пространственной и временной неоднородностью среды, на основе анализа процессов их развития и функционирования.
Объектом изучения послужили основные представители видов-обрастателей, широко распространенные в верхних отделах шельфа. Выбор объекта изучения определялся его спецификой, позволявшей использовать в равной мере как традиционные гидробиологические, так и другие методы. Материал работы составили результаты исследования в теплое время года гидробиологической съемки в трех районах прибрежья г. Махачкалы. Сезонные сборы дали возможность рассмотреть развитие и функционирование сообществ в динамическом аспекте.
Цель работы определила основные задачи исследования:
1) изучить таксономический состав, выявить сезонную и многолетнюю динамику видового состава, видового разнообразия, биомассы зообентосной составляющей, структуры сообщества в связи с обитанием в разных экологических зонах прибрежья;
2) выявить роль и место разных типов обрастания в прибрежной экосистеме;
3) разработать типологию прибрежных сообществ обрастания
Научная новизна. В результате проведенного исследования разработана типология прибрежных сообществ обрастания; предлагается номенклатура и классификация типов прибрежного сообщества обрастания; выявлена трофическая и ярусная структура разных типов сообществ обрастания; исследована сезонная динамика биоразнообразия выделенных типов сообществ; выявлены сезонные и многолетние особенности формирования сообществ разных типов; показано, в верхних зонах шельфа видовое разнообразие закономерно увеличивается от супралиторального к сублиторальному типу сообществ; выявлены сезонные фазы в развитии разных типов сообществ обрастания, показано, что для выявленных типов прибрежных сообществ обрастания характерно разное число фаз; установлено время жизни разных типов сообществ обрастания, время жизни супралиторального сообщества составило от нескольких дней до нескольких недель, псевдолиторального Ч от нескольких месяцев до нескольких лет, существование сублиторального сообщества растягивается на годы.
Теоретическая и практическая значимость. Изучение сукцессии обрастания и установление сроков формирования сообществ имеют большое практическое значение, поскольку позволяют прогнозировать процесс обрастания сооружений и механизмов, эксплуатируемых в морских условиях. Практика эксплуатации судов и гидротехнических сооружений показывает, что отсутствие тщательного изучения экологии обрастателей в конкретных условиях может привести к тому, что меры борьбы с ним окажутся малоэффективными и экономически невыгодными. С другой стороны, исследования обрастания помогут нахождению новых путей повышения продуктивности морской аквакультуры.
Апробация работы. Основное содержание работы отражено в 8 публикациях, из которых 3 в журналах, включенных Структура и объем работы. Диссертация состоит из введения, четырех глав, выводов, библиографического списка и приложений. Работа изложена на 123 страницах, включая приложения, и содержита 18 рисунков, 51 фотографий и 6 таблиц. Список литературы включает 197 наименований, в том числе 53 на иностранном языке.
Благодарность: Автор выражает искреннюю благодарность научному руководителю д.б.н. В.А.Зайко, а также член-корр. РАН д.б.н., профессору М.-Р.Д. Магомедову способствовавшим выполнению настоящей работы советами, критическими замечаниями, представлением материалов. Особую признательность хочу выразить заведующему лабораторией Ботанического Сада ДНЦ РАН Мусаеву А. за предоставленную возможность работы с программой Statistica, StatSoft.
Глава 1. Сообщества обрастания: подходы к их выделению. характеристика основных понятий
В главе рассмотрены существующие представления о понятии сообщества в экологии, основные положения теории развития сообществ, общие взгляды на методологические основы типологии сообществ и проанализированы основные этапы изучения каспийских сообществ обрастания.
Выделение, разграничение и классификация сообществ - одна из центральных проблем современной экологии, тесно связана с исследованием их пространственной структуры, под которой обычно понимают закономерное распределение характеристик богатства и обилия видов в среде обитания. В соответствии с этим выделение морских сообществ (комплексов, трофических зон и т.п.) в большинстве работ проводилось по принципу доминирования определенного вида или группы организмов. Качественным и количественным показателям пространственной структуры морских сообществ разные авторы придают, однако, разное значение. В зависимости от того, чему автор придает решающее значение в формировании сообщества, в гидробиологии сформировалось два основных подхода к выделению сообществ: биотический и фациальный (Кусакин, 1982).
Традиционное выделение сообществ по критериям сходства-различия видового состава и численности особей разных видов, собранных на отдельных гидробиологических станциях, приводит к неоднозначности в интерпретации и даже к явным противоречиям со здравым смыслом, так как позволяет бесконечное дробление сообщества. Как свидетельствует длительная история развития концепций сообщество в экологии, каждый из реализуемых подходов к их выделению обладает определенными преимуществами и богатейшими методическими возможностями. Критический обзор существующих представлений показал, что наиболее надежные критерии для выделения морских сообществ могут быть получены лишь на основании анализа закономерностей их развития и функционирования. Исследование сообщества обрастания рассматривается в работе в качествеа модели морского прибрежного сообщества. Основной анедостаток изучения явления обрастания заключается в том, что в аповседневной практике исследования морских сообществ их характеристики оказываются выхваченными из процесса его развития в период отбора проб. Отсюда причины анализируемого слепка, как правило, остаются не раскрытыми, так как история формирования и закономерности функционирования данного сообщества не известны. Этим, по нашему мнению, определяются многие существующие противоречия интерпретации типов обрастания.
Исследование ранних стадий развития сообществ обрастания достаточно хорошо известны. Однако обобщить и тщательно проанализировать существующий опыт мешает неструктурированное многообразие типов обрастания. Поэтому разобраться в их классификации и типологии - актуальная задача. Разработанная классификация и типология, в свою очередь, должна послужить основанием для проведения долговременных наблюдений за сообществами. аЗдесь важно проследить всю цепочку развития сообщества от зарождения до его полного преобразования. А также объективно склеить аранние стадии развития сообществ с существующими слепками поздних стадий, тем самым решительно продвинуться в понимании этого явления.
Глава II. Физико-географическая характеристика
района и методика исследования
Объектами исследования служили виды макробентоса, составляющие сообщества обрастания, находящиеся в разных экологических зонах моря (супралиторали, псевдолиторали и в верхних отделах сублиторали).
Исследовали формирование сообщества обрастания на разных биотопах - естественных (валунные и скальные гряды) и искусственных (причальные и защитные стенки нового строящегося порта). Пробы отбирали на участках махачкалинского прибрежья (участки: Порт, Камни и Поселок), в каждом из которых обследовали от 5-7 до 18- 20 участков. Учитывали влияние волнового воздействия на сообщество, поэтому в природных экспериментах пробы отбирали в точках, которые былиаа открыты волновому воздействию, а также с полуоткрытых и закрытых участков прибрежья. Для этого в теплое время года (конец мая - октябрь) отбирались пробы раз в 10 - 20 дней. В тех случаях, когда анализировали процесс обрастания биотопа макрофитами, пробы отбирали каждые два дня и чаще. Как правило, синхронно с отбором проб фотографировали сообщество обрастания. Такая детальность отбора проб во времени, как показало наше исследование, позволяет анализировать динамику формирования сообщества. Были просмотрены площадки на камнях (10Х10 см), рассчитывалось среднее значение из двух проб. Попавшие в пробу животные и растения подсчитывали и взвешивали. Методика исследования опубликована (Зайко и др., 2005). Просмотрено около 996 проб, проанализировано 356, сделано более 1500 фотографий.
Для выявления особенностей характера сукцессии определялись процентные доли отдельных таксономических групп организмов в общей биомассе обрастания. С той же целью использовался показатель видового разнообразия Шеннона - Уиллера (Одум, 1975).
n
H = Ч ?pi lnpi; аpi = Bi/B
I =1
где: рi- показатель относительного обилия данной таксономической группы (видового ранга); Вi - биомасса отдельной таксономической труппы; В - общая биомасса обрастания.
Глава III. Сообщество обрастания в разных экологических зонах прибрежья западной части среднего каспия
В проведенном исследовании пытались ответить на вопрос, как развивается и функционирует сообщество обрастания в прибрежной зоне в разных экологических условиях? Для этого в разных прибрежных экологических зонах моря (супралиторали, псевдолиторали, сублиторали) проведен сравнительный анализ видового состава сообщества обрастания; проанализирована сезонная и многолетняя динамика биомассы зообентоса; выявлена сезонная и многолетняя динамика видовой, трофической структуры, биологического разнообразия, оценено время жизни сообщества.
Сравнительный анализ видового состава сообщества обрастания в разных экологических зонах моря позволяет получить общее представление о прибрежном сообществе обрастания и подойти к решению вопроса о реакции сообщества на разные экологические условия его обитания (рис.1). В смысле видового богатства обрастание принято рассматривать как обедненный бентос и даже как крайний случай олигомиксных донных биоценозов (Турпаева, 1977; Брайко, 1985).
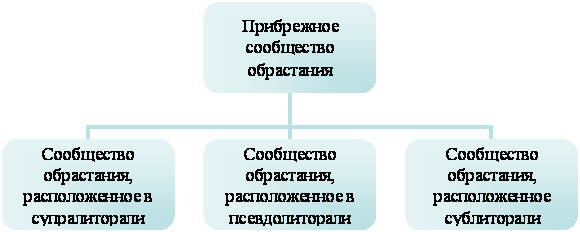
Рис. 1. Схема дифференциации сообщества обрастания по разным экологическим зонам моря, принятая в работе.
Таблица 1.
Видовой состав выделенных типов сообществ обрастания
Макрозообентос |
Типы обрастания |
|||||||
Виды |
1.1.вл |
1.1.оз |
1.2 |
2.1 |
2.2 |
2.3 |
3.1 |
3.2 |
Mytilaster lineatus |
|
|
+ |
+ |
+ |
+ |
+ |
+ |
Balanus improvisus |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Dikerogammarushaemobaphes |
|
|
|
|
+ |
+ |
+ |
+ |
Dikerogammarus sp. |
+ |
|
|
+ |
+ |
+ |
+ |
|
Amathillina cristata |
|
|
|
|
+ |
+ |
|
+ |
Amathillina affinis |
|
|
|
|
+ |
+ |
+ |
|
Niphargoides robustoides |
|
|
|
|
+ |
+ |
|
|
Gammarus placidus |
|
|
|
|
|
|
+ |
|
Pontogammarus maeoticus |
+ |
|
+ |
|
|
|
|
|
Palaemon elegans |
|
|
|
|
+ |
+ |
+ |
+ |
Conopeum seurati |
|
|
|
|
|
|
+ |
+ |
Nereis diversicolor |
|
|
|
|
|
|
+ |
+ |
Mercierella enigmatica |
|
|
|
|
|
|
+ |
+ |
Fabricia sabella ssp. сaspica |
|
|
|
|
|
|
+ |
+ |
Rhitropanopeus harrisii |
|
|
|
|
+ |
+ |
+ |
+ |
Cordylophora caspia |
|
|
|
|
|
|
+ |
|
Dreissena rostriformis |
|
|
|
|
|
|
+ |
+ |
Theodoxus pallasi Ldh |
|
|
+ |
|
|
|
|
|
аPterocuma pectinata |
|
|
|
|
|
|
+ |
|
Schizorhynchus bilamellatus |
|
|
|
|
|
|
+ |
|
Corophium chelicorne |
|
|
|
|
|
|
+ |
+ |
Corophium spinulosum |
|
|
|
|
|
|
+ |
+ |
Где: 1. Супралиторальный тип обрастания (1.1. сезонный (весенне-летний (1.1.вл) и осенне-зимний (1.1.оз), 1.2. Супралиторальных ванн. 2. Псевдолиторальный тип сообщества обрастания (2.1. Квазиэфемерный, 2.2. Псевдолиторальный, 2.3. Псевдолиторальных ванн). 3. Сублиторальный тип обрастания (3.1 Сублиторальный бентосный, 3.2. Сублиторальный пелагический).
Еще в большей степени, как показали наши исследования, это характерно для каспийского прибрежья (супралиторали, псевдолиторали). В результате гидробиологических съемок, проведенных в течение нескольких лет в сообществах обрастания, расположенных в разных экологических зонах моря, регистрировалось от 1-5 до 22 видов (табл.3.1).
По литературным данным, сообщества обрастания в разных частях Каспийского моря имели в своем составе от 8 до 18-ти видов (Зевина, 1982;
Багиров, 1978; Алигаджиев и др., 2001). Эта величина варьирует в зависимости от времени года, свойств доминантов и субдоминантов и условий района в пределах Каспийского моря, в котором находится сообщество (Зевина, 1962). Так, морские сообщества обрастания разнообразнее и богаче в видовом отношении, чем прибрежные (Зевина, 1962; Багиров м др., 1978; Алигаджиев и др. 2001, и др.). По данным В.Кургеняна (1979), общее число видов зоомакробентоса, расположенных рядом с сообществами обрастания естественных экосистем, превышает 24 вида.
Анализ таблицы 1 показывает, что в разных экологических зонах моря видовой состав сообщества меняется. Установлено, что число видов растет с продвижением в ряду - супралиторальные - псевдолиторальные - сублиторальные сообщества.
Фазы развития. Выделены четыре основные фазы сезонного развития сообщества обрастания: начального обрастания - мартЦначало апреля (1); роста - апрель - конец мая (2); зрелости - июнь - середина сентября (3); и переходного периода - середина сентября - февраль (4).
Развитие зообентосной составляющей рассмотрим на примере сообщества, расположенного в псевдолиторальной зоне шельфа. Первая фаза сукцессии в зависимости от времени года, локальных экологических особенностей местообитания может иметь несколько вариантов сообществ: водорослей, митилястера, балянуса или различных их сочетаний. Вторая фаза - роста. Биомасса зообентосной составляющей (сообществ митилястера и балянуса) растет исключительно за счет переживших зиму особей. Этим объясняется пологий участок на кривой динамики биомасс. Рост биомассы в этом временном промежутке в основном происходит в результате роста биомассы макрофитов (на схеме биомасса макрофитов не учитывалась). С середины лета (в зрелой фазе), когда в сообществе появляются вагильные формы (гаммариды, креветки, крабы), доля зообентосной составляющей резко возрастает. Кроме того, в эту фазу наблюдаются, как правило, несколько волн активного оседания усоногих раков. Об этом свидетельствует наличие разноразмерных домиков балянусов на псевдолиторали. Постепенно со временем участки оседания балянусов в верхней части псевдолиторали зарастают полностью. В конце зрелой фазы, приблизительно с середины августа, балянусы начинают оседать вторым слоем на ранее осевших усоногих раков, многие из которых, лишившиеся возможности существовать, отмирают.
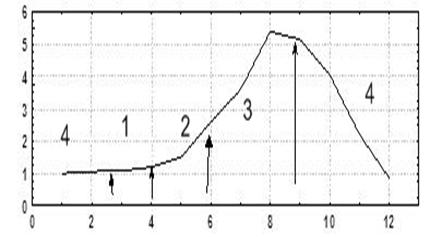
Рис. 2. Схема фаз развития в сезоне в псевдолиторальной зоне шельфа зообентосного сообщества обрастания.
По оси абсцисс - время, месяцы года;
По оси ординат - биомасса зообентосной составляющей, условные единицы.
Во втором слое можно встретить и раковины митилястера, и талломы макрофитов. Затем, в связи с началом периода осенних штормов, наблюдается резкое падение температура воды прибрежной зоны моря. При этом биомасса сообщества начинает снижаться. С ее падением вагильные формы покидают прибрежье, уходя на глубину (Гусейнов, 2005). Это приводит к еще аболее резкому падению биомассы зообентоса (рис. 2). Таким образом, с наступлением фазы переходного периода (осенью) происходит резкое падение биомассы зообентосной составляющей. Эта тенденция прослеживается и зимой вплоть до конца февраля, начала марта следующего года, однако, скорость снижения биомассы в этот период значительно меньше.
На псевдолиторали и сублиторали сообщество обрастания проходит все четыре сезонные фазы, которые затем в процессе развития сообщества многократно повторяются. Часть сообщества обрастания, находящаяся в пределах псевдолиторали в зоне прибоя, названная нами эфемерной, как правило, новый сезон начинает с чистого субстрата. Сообщество обрастания, находящееся в псевдолиторали вне зоны прибоя, как и сублиторали, начинают новый сезон с некоторой минимальной биомассы, которая осталась от предыдущего года. По-видимому, весенняя биомасса сообщества обрастания определяется складывающимися условиями в этих экологических зонах в предшествующий зимний период.
Продолжительность жизни сообщества. Следует обратить внимание, что использование этой характеристики имеет важный общетеоретический интерес, так как о стабильности и устойчивости сообщества судят, как правило, с учетом этого фактора. Наши данные показали, что время жизни в разных экологических условиях прибрежных сообществ обрастания различно (Имашова, Зайко, 2010; Имашова, 2010).
Для супралиторального сообщества, живущего относительно короткий период в течение менее чем одного года (сезона) и названного нами фантомным, характерно возрождение в разные сезоны года, в разных частях супралиторали, в разных сочетаниях видов.
Если продолжительность жизни эфемерного псевдолиторального сообщества, расположенного в пределах зоны прибоя, составляет время меньше чем один сезон, то новый сезон начинается с нулевой биомассы. Если в условиях прибрежья ему удалось пережить зиму, то весной сообщество начинает с некоторой биомассы, и стадии сезонной сукцессии во второй год жизни повторяют стадии развития характерные для вечных квазиэфемерного псевдолиторального и сублиторального обрастания.
С учетом биологического времени жизни сообщества (учитывая число фаз его развития) можно дать и другую оценку продолжительности его жизни. Фантомное аразвитие охватывает время от части одной фазы, до нескольких, и, как правило, это сообщество не проходит все четыре сезонные фазы развития. Развитие эфемерного и квазиэфемерного сообщества, расположенных в псевдолиторали, - захватывает все четыре фазы сезонного развития. Кроме того, для вечных типов и подтипов сообществ обрастания характерно многократное повторение сезонных фаз развития.
Таблица 2.
Общая статистическая модель
Зависимая переменная |
Множественный R |
Множественный R2 |
F |
p |
V8 |
0,659630 |
0,435112 |
8,216118 |
0,000340 |
V11 |
0,597751 |
0,357306 |
5,930134 |
0,002458 |
V14 |
0,559879 |
0,313465 |
4,870294 |
0,006682 |
аГде: V8 - биомасса зообентоса весной, V11 - биомасса зообентоса летом и V14 - биомасса зообентоса осенью.
Мы экспериментально оценили время жизни супралиторального сообщества обрастания. Наши наблюдения показали, что ширина занимаемой зоны и время существования сообщества зависят от многих факторов. Во-первых, от степени защищенности исследуемого участка от волнового воздействия (открытые, полуоткрытые и закрытые участки). Согласно нашим оценкам средняя биомасса супралиторального сообщества на дагестанском прибрежье Среднего Каспия меняется в зависимости от места расположения исследованного участка. В закрытых участках она достигает значений до 10 г/м2, в полуоткрытых рядом с зоной прибоя - до 25-30 г/м2 . На открытых относительно пологих участках в прибойной зоне после летнего продолжительного шторма может возрастать на порядок (до 250-300 г/м2). Во-вторых, от продолжительности и интенсивности прибоя. В-третьих, от положения субстрата относительно плоскости моря (вертикальная стенка, пологий берег). И, наконец, от особенностей адаптаций животных и растений, входящих в состав сообщества, их способности оставаться жизнеспособными в условиях без постоянного контакта с морской водой.
Результаты анализа сведены в таблице 2. Проведенный анализ апоказал (Имашова, 2010), что процессы, происходящие в скалистом сообществе обрастания, расположенном в супралиторали в прибрежье Махачкалы, описываются статистической моделью (включающей все независимые факторы, описанные выше) с высокой степенью достоверности (р < 0.001). Эти факторы на 93% определяют изменчивость времени жизни сообщества.
Таким образом, время жизни в разных экологических условиях прибрежных сообществ обрастания различно. Если учитывать время жизни разных сообществ, то можно выделить следующие типы сообществ: фантомное, лэфемерное, квазиэфемерное и вечное. Кроме того, существует и другой способ оценки продолжительности жизни, с учетом биологического времени сообщества, - по числу фаз развития. Тогда эфемерный тип развития охватывает время от части одной фазы до нескольких. Как правило, это сообщество не проходит все четыре выделенные нами фазы сезонного развития. Развитие квазиэфемерного псевдолиторального типа сообщества уже охватывает все четыре фазы. Для вечных типов и подтипов сообществ обрастания характерно многократное повторение сезонных фаз развития.
Трофические группировки зообентоса в разных экологических зонах прибрежья.
Проведенный в работе сравнительный анализ сезонной и многолетней динамики видовой, трофической структуры, биологического разнообразия позволил оценить специфику реакции сообщества на закономерно меняющиеся из года в год сезонные и межгодовые колебания внешних условий и дал возможность говорить об общей реакции сообщества на воздействие среды обитания с большей определенностью, чем раньше.
В работе исследовали, как меняется трофическая структура в разных экологических зонах моря в пределах прибрежного сообщества обрастания. Было установлено, что существует вполне отчетливая динамика изменения основных составляющих в зависимости от экологических условий. Доля автотрофов в трофической структуре максимальна в псевдолиторали, вверх от нее к супралиторали и вниз по шельфу к сублиторали биомасса макрофитов резко падает. Доля макрофитов в обрастании прибрежья в теплое время года подавляющая.
Для каждой экологической зоны моря существует вполне определенная трофическая структура (рис. а3, 4). Основной трофической группой зообентоса априбрежного обрастания являются сестонофаги, обитающие на поверхности твердых грунтов (СПО - балянус и митилястер). аИх доля в обрастании меняется в сезоне, что связано с сезонностью их размножения и спецификой роста моллюсков. Выявлены также межгодовые колебания биомассы всего сообщества в пределах от 23.5% до 89.23%.
Гаммариды и кумовые - собирающие детритофаги, обитающие на поверхности осадков (СДПГ). Они практически отсутствуют в супралиторали и представлены в псевдолиторали и сублиторали. Из этой трофической группы - гаммариды в псевдолиторали достигают наибольшего представительства. Их доля в трофической структуре сообщества обрастания может достигать 25-37%.
Нереис - типичный представитель собирающих детритофагов, обитающих в толще грунта (СДТГ), появился в обрастании в зоне сублиторали лишь в последние годы наблюдений, только после того как в пределах причальной и защитных стенок порта в результате осадконакопления стал формироваться биотоп мягких грунтов.
Плотоядные в прибрежных водах представлены в первую очередь крабом (Rhitropanopeus harrisii), в районе строящегося порта он был отмечен лишь на 3-4 год с начала строительства причальной стенки и до настоящего времени широко не представлен в фауне строящегося морского порта. Доля плотоядных максимальна в зоне сублиторали, однако, ее величина не превышает 7,3%.
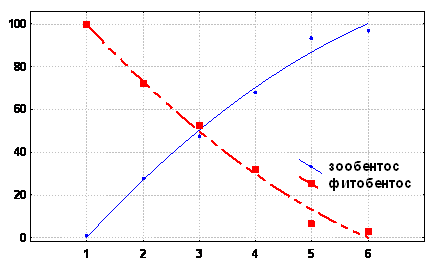
Рис. 3 Основные составляющие прибрежного биоценоза в разных экологических зонах моря (1 - супралитораль, 2 - участки открытой псевдолиторали, 3 - участки закрытой псевдолиторали, 4 - сублитораль бентосная, 5 - сублитораль пелагическая (корабль 6 - сублитораль пелагическая (буи)).
По оси абсцисс: экологические зоны моря, 1- 6. По оси ординат: доля зообентосной и фитобентосной, составляющих в разных экологических зонах моря, % .
Детритофаги, питающиеся из толщи грунта (ДТГ), в сборах отсутствовали.
В исследованных экологических зонах моря представительство разных трофических групп зообентоса меняется. Отмечена тенденция уменьшения количества групп при переходе от сублиторали (5 трофических групп) к псевдолиторали и супралиторали (по 3 трофических группировки) рис. 4. Доля автотрофов в трофической структуре максимальна в псевдолиторали, вверх от нее по шельфу к супралиторали и вниз к сублиторали биомасса макрофитов резко падает. В общем случае доля отдельных составляющих трофической структуры меняется в сезоне, что связано с выеданием их рыбами-бентофагами, сезонным характером размножения, развития и роста разных групп макробентоса.
На супралиторали она наиболее проста и для фантомных сообществ в зависимости от складывающихся условий в разные сезоны года может включать один или несколько видов обрастателей. Эти особенности структуры во многом объясняются относительно упрощенной ярусностью сообщества обрастания. В результате малого числа видов, успешно адаптированных к экстремальным условиям супралиторали, трофическая структура может состоять из одного - двух видов, при доминировании
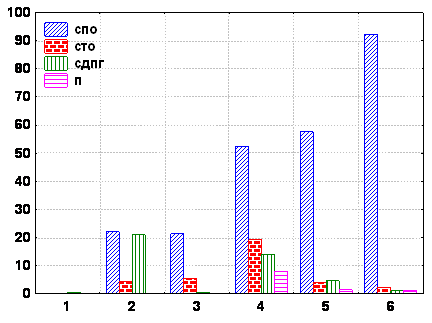
Рис. 4 Состав трофических группировок зообентоса в разных экологических зонах моря
По оси абсцисс: экологические зоны моря, цифры с 1 до 6
(обозначения то же, что и на рис. 3).
По оси ординат: доля трофической группировки в разных экологических зонах моря - а%.
макрофитов (автотрофов). Если поселение балянуса и митилястера на твердых субстратах рассматривать как биотоп, то он обладает своеобразной двойственностью. С одной стороны, убежищем и местом прикрепления животных служат непосредственно камни и скалы, а с другой, - друзы и поверхность раковин животных. Следствием этого и оказывается наблюдаемая ярусность в пространственной структуре донного сообщества.
Таким образом, трофическая структура исследуемых сообществ обрастания в разных экологических зонах прибрежья меняется, отражая специфику условий обитания. Проведенный анализ показал, что соотношение автотрофов и гетеротрофов закономерно уменьшается при переходе от супралиторали к псевдолиторали и сублиторали. Для числа трофических группировок отмечена противоположная тенденция.
В зоне псевдолиторали и сублиторали сложность ярусной структуры сообщества возрастает, в результате создаются условия, которые благоприятны вагильным видам (черви, крабы, креветки, гаммариды). Для псевдолиторали и сублиторали она наиболее сложна, в ее состав входит максимальное число трофических группировок. В исследованных экологических зонах моря представительство разных трофических групп зообентоса меняется. Отмечена тенденция уменьшения количества групп при переходе от сублиторали (5 трофических групп) к псевдолиторали и супралиторали (по 3 трофических группировки).
Видовое разнообразие. Видовое разнообразие оценивали по формуле Шенонна-Уиллера. Как показали наши исследования, на разных участках псевдолиторальных прибрежных сообществ обрастания видовое разнообразие в сезоне растет от зимы к осени и затем закономерно снижается к зиме (Зайко и др., 2006). Биоразнообразие сообщества обрастания, расположенного в порту в пределах сублиторали, со временем постепенно возрастало и в основном за счет появления вагильных видов, а также видов, атребующих особых условий для своего существования. Например, нереис появился на третий - четвертый год, когда в результате формирования ярусной структуры сообщества создались условия для успешного осадконакопления. Нами было отмечено, что в пределах участков, занятых сообществом обрастания, сформирован новый биотоп - тонкий слой мягких грунтов, столь необходимый для существования нереиса.
Глава IV. Типология прибрежного
сообщества обрастания
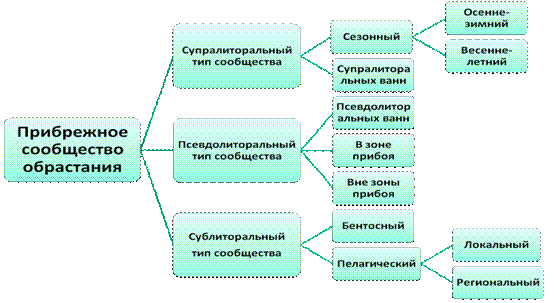
В соответствии с подходом, развиваемым в данном исследовании, были проанализированы основные характеристики сезонной и многолетней динамики выделенных типов сообщества обрастания. Как показали наши наблюдения за прибрежными типами сообществ обрастания, эти характеристики определяются как неоднородностью экологических условий, которые складываются на прибрежье,а так и сезонностью в размножении, успешностью в оседании и развитии составляющих сообщество видов. В результате была разработана схема иерархической структуры прибрежного сообщества, которая представлена на рис.5. Результаты исследования позволяют сделать вывод, что выделенные типы сообщества обрастания отличаются по основным характеристикам, используемым для выделения сообществ - числом видов, их сезонной динамикой, временем жизни, трофической структурой, видовым разнообразием, ролью в функционировании прибрежной экосистемы. Кроме того, детальный анализ функционирования отдельных типов сообщества обрастания выявил различия в сезонном функционировании отдельных типов сообществ, в зависимости от складывающихся условий и особенностей адаптаций видов, их составляющих, и поставил вопрос о существовании подтипов этих сообществ.
1. Супралиторальное сообщество обрастания (лфантомное)
Супралиторальная зона, расположена на шельфе выше литорали, может заливаться водой лишь спорадически, при нагонах или сильных штормах. В остальное же время супралитораль нерегулярно смачивается брызгами прибоя. Число видов животных и растений, способных существовать в этих суровых условиях между сушей и морем, невелико, возможно, что эта простота сообществ определяет то, что супралиторали уделяется гораздо меньшее внимание, чем, например, скалистой литорали и сублиторали (Кусакин, 1977; Ошурков, 1987; Menge, Branch, 2001; Menge, Sutherland 1987, и мн.др.).
а
Рис. 5 аСхема предлагаемой иерархической структуры
прибрежного сообщества обрастания
Видовое разнообразие скалистого супралиторального сообщества весьма невелико. Список видов этого типа сообщества обрастания представлен в таблице 1. Для супралиторального сообщества, живущего относительно короткий период, в течение менее чем одного года (сезона), и названного нами фантомным, характерно возрождение в разные сезоны года, в разных частях супралиторали, в разных сочетаниях, составляющих его видов.
Выявлены основные факторы, влияющие на время существования этого типа сообщества (Имашова, 2010). Проведенный анализ показал, что с высокой степенью достоверности (р < 0.001) и выбранные нами факторы: степень защищенности исследуемого участка от волнового воздействия (открытые, полуоткрытые и закрытые участки; продолжительность и интенсивности прибоя; положения субстрата относительно плоскости моря (вертикальная стенка, пологий берег) - на 93% определяют изменчивость времени жизни сообщества.
1.1. Супралиторальный сезонный подтип
В зоне заплеска были выделены две формы сообществ - осенне-зимняя и весенне-летняя, что связано, по-видимому, в первую очередь с существующей сезонностью оседания личинок и спор. Оба подтипа сообщества имели разный видовой состав, но сходные фазы, развития и деградации сообщества - организмов обрастателей (Зайко и др., 2006).
1.1.1 Сезонный весенне-летний. Список видов этого типа обрастания представлен в таблице 3.1. Продолжительность существования этого подтипа составила в разных районах западной части Среднего Каспия разное время в прибрежье г. Махачкала (2-4 недели), в его состав, в отличие от осенне-зимней формы, входили два вида зеленых водорослей и гаммариды. На юге района исследования продолжительность жизни этого подтипа сообщества обрастания еще больше сокращалась - до 5-12 дней (Имашова, 2010). Гибель этого подтипа сообщества была обусловлена штилевой и жаркой погодой. По-видимому, отмеченные различия в продолжительности жизни подтипов фантомных сообществ в большей мере обрастания объясняются сезонной спецификой климатических и гидрологических условий в этих районах.
1.1.2. Сезонный осенне-зимний. В состав осенне-зимнего эфемерного сообщества обрастания входили лишь макрофиты (красные и зеленые водоросли). Время существования этого сообщества от зарождения до исчезновения составило около двух месяцев. Удалось детально зафиксировать с помощью цифрового фотоаппарат развитие, деградацию и гибель сообщества в 2005 г. в районе махачкалинского района Камни После непродолжительного развития сообщества, приблизительно в течение месяца, сначала его засыпало песком после сильного шторма, когда песок через две недели смыло в море, остатки сообщества покрыло снегом и льдом. Когда лед растаял, стало видно, что эфемерное сообщество обрастания погибло.
1.2. Супралиторальных ванн. Другой разновидностью супралиторального типа обрастания служат сообщества супралиторальных ванн. Для этого подтипа характерны резкие колебания условий среды обитания. После сильных дождей вода иногда почти полностью опресняется, в жаркую погоду, наоборот, соленость ее может превышать обычную морскую. Температурные колебания также весьма велики. Фауна и флора этих ванн содержит небольшое число эврибионтных морских видов. Как правило, им свойственна низкая продуктивность. Что, в первую очередь, определяется низкой численностью животных и растений, широкими колебаниями температуры и солености. Продолжительность жизни этого подтипа сообщества зависит в первую очередь от сгонно-нагонных явлений и подвижности песчаного субстрата.
Таким образом, супралиторальный тип сообщества обрастания - это маловидовое, физически контролируемое сообщество с относительно коротким временем жизни, в асостав которого входят животные и макрофиты с исключительными адаптациями к обезвоживанию и пересыханию (Имашова, 2010, Имашова, Зайко, 2010).
2. Псевдолиторальное сообщество обрастания. Протяженность псевдолиторали по вертикали весьма невелика (несколько десятков сантиметров), но при очень пологих берегах ширина может достигать нескольких метров. И здесь, как на супралиторали, резко выражены суточные и сезонные колебания температуры, солености, увлажнения и другие.
Развитие процесса сукцессии сообщества обрастания в прибойной зоне описано нами в предыдущей главе. Первая фаза сукцессии в зависимости от времени года, локальных особенностей биотопа может иметь несколько вариантов: сообщество водорослей (Enteromorpha spp.); сообщество митилястера - (Mytilasterlineatus); сообщество балянусов - (Balanus improvisus). Первоначальные отдельные пятна балянусов постепенно в течение лета превращаются в почти сплошную щетку, особенно в той части валунов, которые подвергаются волновому воздействию. Характер распределения обрастания рачками носит диффузный характер, с максимальной плотностью особей вдоль линии уровня моря, что хорошо видно во время штиля. К лету водоросли начинают доминировать по биомассе. С этого времени их влияние на зообентосную составляющую сообщества обрастания значительно. Отмечен четко выраженный сезонный характер изменения численности и биомассы гаммарид. Максимальная численность и биомасса бокоплавов отмечена в период с середины июля до середины августа. Роль макрофитов для подвижных форм во многом объясняется тем, что они используются ими как укрытие, как один из основных компонентов пищи. С развитием макрофитов растет биологическое разнообразие ценоза за счет увеличения подвижных форм зообентоса (гаммариды, креветки). Наибольшее разнообразие достигается в конце лета. Окончание вегетационного периода макрофитов приводит к распаду зообентосного сообщества, остатки которого уничтожаются штормами в осенне-зимний период года.
2.1. Псевдолиторальный, находящийся в зоне прибоя (лэфемерный). Список видов этого типа сообщества обрастания представлен в таблице 1. Развитие процесса сукцессии псевдолиторального сообщества обрастания в прибойной зоне происходит по следующей схеме. Весной, в зависимости от складывающихся условий прошедшей зимы, прежде всего продолжительности и силы штормов, формирование обрастания на разных участках начинается с лостатков биоценоза предшествующего года, или с голого субстрата (Зайко, Амаева, 2005, Зайко и др., 2005). Проведен анализ факторов, влияющих на биомассу зообентоса этого подтипа псевдолиторального сообщества (Имашова, Зайко, 2010). Для того чтобы оценить действие волнового фактора на прибрежные сообщества, участки, выбранные для анализа, сводили в две группы: в пределах зоны прибоя, авне ее. Это было необходимо сделать, так как состав и структура прибрежного квазиэфемерного сообщества в зависимости от размещения сообщества относительно зоны прибоя отличались (Зайко и др., 2006). При реализации статистической модели в качестве группирующих были выбраны следующие факторы: годы (условное обозначение - V2) - фактор, отражающий сезонную динамику характеристик сообщества за разные годы наблюдений с 2004 по 2007 г; лучасток (V3) - районы сбора зообентоса: порт, камни, поселок; воздействие (V5) - участки с разной степенью волнового воздействия: открытые, полуоткрытые, закрытые (лоткрытые - участки прямого волнового воздействия, полуоткрытые - находящиеся внутри порта, на защитных стенках с внешней стороны; закрытые - находящиеся внутри порта за защитными стенками с внутренней стороны). В качестве независимой переменной выступала биомасса зообентоса за весенний (V8), летний (V11) и осенний (V14) сезоны. Результаты анализа сведены в таблице 3. Анализ таблицы показывает, что модели для всех зависимых переменных описывают процессы, происходящие в сообществах обрастания в прибрежье Махачкалы, с высокой степенью достоверности (р < 0,01). Выбранные ав модели независимые факторы: участок исследования (лпорт, камни, поселок), астепень открытости волновому воздействию (открытые, полуоткрытые, закрытые) - оказывают влияние на биомассу зообентоса наиболее полно весной (43%).
Таблица 3
Общая статистическая модель
Зависимая переменная |
Множественный R |
Множественный R2 |
F |
p |
V8 |
0,659630 |
0,435112 |
8,216118 |
0,000340 |
V11 |
0,597751 |
0,357306 |
5,930134 |
0,002458 |
V14 |
0,559879 |
0,313465 |
4,870294 |
0,006682 |
Это объясняется тем обстоятельством, что весной зообентос представляют в основном усоногие раки и митилястер, пережившие зиму, и их биомасса во многом определяется условиями, сложившимися в предшествующий осенне-зимний сезон и в первую очередь - степенью и продолжительностью волнового воздействия (Имашова, Зайко, 2009).
Как известно, в летний сезон у особей усоного рака и двустворчатых моллюсков отмечается активный рост. Биомасса зообентоса при этом испытывает разнонаправленное воздействие. С одной стороны, численность макрозообентоса сокращается вследствие хищничества. С другой - происходит активный рост предыдущих генераций сестонофагов и вновь осевшей молоди. Преобладание того или иного фактора в разных местах сбора животных меняет биомассу зообентоса. В модели такое влияние учитывалось лишь косвенно. Это, по-видимому, и объясняет снижение влияния волнового фактора на биомассу в летний период наблюдений за сообществом до 36%.
Осенью к выше описанным факторам добавляется еще один, связанный с особенностями оседания балянусов - формированием и разрушением сообщества обрастания, имеющего многослойную структуру (Имашова, Зайко, 2009). Действие этого фактора меняется как во времени, так и в пространстве. Как правило, с формированием многослойного сообщества сначала происходит небольшое увеличение биомассы бентоса, а затем со временем она уменьшается. На биомассу зообентоса оказывает влияние проросшие в ней макрофиты. После достижения ими некоторого определенного размера, даже при небольшом волнении моря, в пределах обрастаний, которые имеют многослойную структуру, ее часть может быть оторвана от биотопа, и на нем образуются пустые места без обрастания (лплешины). И этот явно неучтенный в нашей модели фактор, по-видимому, еще больше снижает оценку влияния волнового фактора на биомассу зообентоса в осенний период (до 31%). Так как готовность к разрушению многослойной структуры, как мы уже отмечали, зависит не только от степени волнового воздействия, но и от длины (точнее парусности) проросших в ней макрофитов.
Анализ статмодели показал, что выбранные в качестве независимых факторы: лучасток исследования, степень открытости волновому воздействию, с высокой степенью достоверности (р < 0,01) оказывают влияние на биомассу зообентоса наиболее полно весной (43%), снижаясь соответственно до 36 и 31% летом и осенью. Это послужило основанием для разделения этих двух подтипов псевдолиторального сообщества обрастания.
2.2. Псевдолиторальный, находящийся вне зоны прибоя (лквазиэфемерный). Список видов этого подтипа сообщества обрастания представлен в таблице 1. Он расположен на некотором удалении (5-20 м) от зоны прибоя в зависимости от закрытости берега от прямого волнового воздействия. По-видимому, это уже сообщество с неограниченным временем жизни. В районе строящегося порта в зиму уходят абалянус, митилястер, зеленые и красные водоросли. Со временем биоразнообразие сообщества этого подтипа росло. Если креветки присутствуют уже в первый год формирования сообщества, то первые крабы были обнаружены лишь на четвертый год существования этого типа обрастания.
Таким образом, в отличие от предыдущего подтипа псевдолиторального сообщества обрастания этот уже имеет атрибуты самоподдерживающегося сообщества - у составляющих его видов появляется размерно-возрастная структура популяции. Тем не менее, по-видимому, его благополучное существование аво многом еще зависит от расположенных глубже по шельфу бентосных сообществ-доноров. а
2.3. Подтип псевдолиторальные ванны
В основном в этом подтипе нами отмечены макрофиты, балянус и редко митилястер. В защищенных от волн местах это сообщество высокопродуктивно, в частности биомасса макрофитов может достигать нескольких килограмм на квадратный метр. В незащищенных от волнового воздействия районах, как и в случае с супралиторальными ваннами, основной фактор, контролирующий сезонную динамику сообщества, - волновое воздействие. На прибрежье валунные гряды окружены, как правило, песчаным субстратом. В период штормов ванны то засыпаются, то очищаются от песка.
3. Сублиторальный тип обрастания. а
Важность проведения исследований сублиторальных типов обрастания определяется многими причинами. В первую очередь, ролью этого типа сообщества в прибрежной экосистеме (Witman и Sebens, 1992; Dugan и Davis 1993; Dayton et. al., 1995 и мн.др.). Во-вторых, для преодоления нерешенных теоретических и практических проблем, в частности, накопившихся в результате отсутствия критериев адекватного сравнения пелагических и бентосных сообществ обрастания (Ошурков, 1985, 2000 и др.). Кроме того, полагают, что устранение большинства экологических угроз морскому биологическому разнообразию, рыболовству и сохранению морских резерватов произойдет в средах обитания, которые расположены по шельфу ниже приливно-отливной зоны (Polis. et. al., 1997;Witman et. al., 2000).
Нами установлено, что формирование этого типа обрастания, расположенного в пределах сублиторали, на защитной стенке порта, происходит по описанной выше схеме квазиэфемерного типа. К началу зимнего сезона в состав этого сообщество входят, как правило, макрофиты, балянус и митилястер. За весь период наблюдений это сообщество обрастания продолжало существовать, испытывая регулярные сезонные изменения численности и биомассы составляющих его видов.
В зависимости от расположения сообщества в водной толще, нами выделены два подтипа: бентосное и пелагическое сообщества обрастания.
3.1 Подтип бентосное сообщество обрастания. Согласно нашим наблюдениям этот тип обрастания обнаружен в порту на глубинах около 2 м на некотором удалении от зоны прибоя. Его состав и структура весьма сходны с сообществами обрастания, описанными ранее для Каспийского моря (Зевина и др., 1963, Зевина, 1972, 1994 Багиров, 1968, 2000 и др.). При сравнительном анализе материалов по каспийскому обрастанию обнаруживается, что число и состав видов, присутствующих в обрастании более 50 лет назад и теперь, близки. Это свидетельствует о большей устойчивости этого типа сообществ в сравнении с супралиторальным и псевдолиторальным типами сообществ.
3.2. Подтип пелагическое сообщество обрастания
Для преодоления проблем, связанных с типологией сообществ обрастания Резниченко О.Г. была предложена биотопическая классификация обрастания. Для этого он объединил основные виды и группы субстратов обрастания в семь типов (Резниченко, 1978). Нами исследовались, согласно предложенной им классификации, в основном подвижные и неподвижные субстраты.
3.2.1. Пелагическое сообщество обрастания (неподвижные субстраты). Анализ пелагических сообществ проводили с использованием тех же критериев, что и бентосных. Основное внимание в работе было уделено их сезонной и многолетней динамике видового состава и структуры сообщества.
Распределение и видовой состав пелагического сообщества обрастания буев (неподвижные субстраты). При сравнении полученных нами данных с результатами предшествующих исследований установлено, что в обрастании буев в разных частях Каспийского моря встречаются практически одни и те же виды организмов-обрастателей. Отличия наблюдаются в относительном обилии разных групп оброста. В большинстве исследованных районов доминируют два вида: усоногий рак Balanusimprovisusи двустворчатый моллюск Mytilasterlineatus. Полагают, что основное значение в этих случаях играет ток воды, омывающий обрастания (Зевина, 1972).
B. improvisusдоминирует в сообществах обрастания на западном прибрежье Среднего и Южного Каспия, восточном прибрежье Среднего и Южного Каспия, другой вид усоногих раков B. eburneus представлен в сообществе обрастания только в Красноводском заливе. Mytilasterlineatusшироко представлен в тех же районах, где и усоногий рак B. improvisus.
Зональность в обрастании буев. Уже первый взгляд на обрастание буев позволяет сделать вывод, что распределение животных на корпусе буя - смешанное (Имашова, Зайко, 2011). В верхней части оброста присутствуют две четкие зоны - зона водорослевой каймы и зона сестонофагов. Ниже этих зон конфигурация распределения зообентоса несколько иная. На корпусе буя уже не образуется зон сплошного оброста. Распределение животных и растений становится мозаичным. Зона водорослевой каймы, ширина которой в разные сезоны года меняется, испытывая закономерные колебания, от весны к лету расширяется, к осени и зиме - сужается и даже может временно исчезать. Следует заметить, что такая же кайма присутствует и в обрастании псевдолиторали и в обрастании судов.
Ниже зоны водорослей - зона сестонофагов. Ее основные представители - усоногий рак B. improvisusи двустворчатый моллюск M. lineatus. Ширина этой зоны, как правило, 30-50 см, особенно отчетливо проявляется в виде белой полосы во второй половине лета, в период массового оседания рачка. Мы отмечаем определенное сходство в распределении обрастателей по поверхности буя и корпусу судна, работающего в пределах акватории махачкалинского морского порта (Имашова, Зайко, 2011г).
Трофическая структура сообщества обрастания буев. В пределах махачкалинского порта в обрастании буев в летнее время доминируют балянус и митилястер - сестонофаги, обитающие на поверхности субстрата (СПО). Их доля превышает 56% общей биомассы. Вторые по биомассе - макрофиты (35%) У сопутствующих видов, составляющих две трофические группировки: собирающие детритофаги, обитающие на понверхности субстрата (СДПГ) - гаммариды и кумовые, доля которых значительно ниже - 0,8. Плотоядные (П) в обрастании представлены в первую очередь крабом (Rhitropanopeus harrisii) и их доля в общей биомассе оброста буев невелика и составляет около 2 %.
Установлено, что в разных районах Каспия при сходном видовом составе обрастания буев существуют и определенные отличия, которые проявляются в специфике трофической структуры сообщества. С продвижением на юг в западной и восточной частях Каспия растет доля сестонофагов и падают доли всех остальных трофических группировок, в первую очередь за счет снижения доли макрофитов и доли детритофагов, собирающих из толщи грунта.
3.2.2. Пелагическое сообщество обрастания (подвижные субстраты). Основное внимание в работе было уделено анализу основных групп животных, встречающихся в обрастании судов с разным эксплуатационным режимом работы.
Трофическая структура сообщества обрастания судов. Как это видно на рис. 3.5. в пределах махачкалинского порта трофическая структура близка к структуре сообщества обрастания буев.
Зональность в обрастании судов. Уже первый взгляд на обрастание корпуса судна позволяет сделать вывод, что распределение животных на корпусе буя - зональное (Имашова, Зайко, 2011). Самая верхняя зона обрастания как и на буе - зона водорослевой каймы, ширина которой в разные сезоны года меняется, испытывая закономерные изменения: весной и летом расширяется к осени и зиме - снижается и даже может временно исчезать. Ниже зоны водорослей - зона сестонофагов. Это в первую очередь усоногий рак B. improvisusи двустворчатый моллюск M. lineatus. Ширина этой зоны, как правило, 30-50 см. Ниже этой зоны конфигурация распределения зообентоса несколько иная. В этой части корпуса уже не образуется зон сплошного оброста (Имашова, Зайко, 2011).
Суда, имеющие частые стоянки, но работающие в порту или ближайшей к нему акватории моря, в распределении обрастателей имеют свои особенности. У них проявляются элементы и горизонтальной зональности. Нос и скуловая части таких судов имеют мозаично расположенные группировки обрастателей, тогда как мидель и участки кормы заселены обростом почти полностью (Имашова, Зайко, 2010).
В общем случае наши исследования подтвердили существующие представления в распределении оброста на судах, стоящих не причале, и судах с районами работ, охватывающих порт и ближайшую к нему акваторию моря (Тарасов, 1962; Рудякова, 1967; Зевина, 1972, Звягинцев, 1980, 1985; Багиров, 1968, 1977; Багиров и др. 1983 и многие другие).
Таким образом, нами в обрастании судов выявлена четкая вертикальная (от ватерлинии вниз к килю) и горизонтальная (от носа до кормы судна) зональность в распределении животных и растений по корпусу судов разного режима плавания. Проведенный нами анализ показал, что стоящие на приколе суда, и суда, работающие в пределах порта (залива), со временем получают сообщество обрастания подобное рядом расположенным гидротехническим сооружениям (буям, защитным стенкам порта и т.д.). Суда, перемещающиеся из одного порта (с присущим ему локальным сообществом обрастания) в другой (из одного региона в другой), имеют сообщество обрастания, которое находится в процессе перестройки от одного видового состава оброста (порт отправления) к другому (порт назначения). Преобразования тем больше, чем больше различаются эти локальные сообщества обрастания. Это послужило основанием в пелагическом сообществе обрастания (подвижные субстраты) выделить два подтипа: локальный (состав видов которого близок к локальному пелагическому сообществу обрастания определенного порта) и региональный (находящийся в процессе перестройки от одного видового состава оброста порт отправления к другому составу видов порт назначения).
Выводы
1. Исследовалось сообщество обрастания зообентоса прибрежья западной части Среднего Каспия. Показано, что в разных экологических зонах моря реакция сообщества обрастания на сезонные и многолетние колебания условий в процессе его функционирования и развития различна. В результате в разных частях биотопа закономерно меняется видовой состав,а структура и продолжительность жизни сообщества. Общий список видов в сообществе обрастания меняется от 1-5 видов макробентоса на супралиторали до 12- 18 видов в сублиторали.
Трофическую структура прибрежного сообщества обрастания составляют в основном четыре трофические группы зообентоса (сестонофаги, обитающие на поверхности твердых грунтов; собирающие детритофаги, обитающие на поверхности осадков; плотоядные; собирающие детритофаги, обитающие в толще грунта). Число трофических группировок растет в ряду выделенных типов сообществ от супралиторального (1-2) к псевдолиторальному и сублиторальному Ч по 4 трофические группировки.
Установлена фазность в сезонном развитии выделенных типов сообществ обрастания. Супралиторальный (лфантомный) проходит в своем развитии от части одной фазы до двух фаз. Эфемерный псевдолиторальный - до 3-4. Псевдоэфемерному и сублиторальным типам сообщества (по нашей классификации - вечным), как правило, не только присущи все четыре фазы сезонного развития, но и апроявления фазности отмечаются в течение последующих лет жизни.
2. Супралиторальный и псевдолиторальный типы сообщества обрастания в большей степени контролируются внешними факторами (т.е. относятся к физически контролируемым сообществам), тогда как сублиторальный тип - биотическим фактором. Тем не менее, как показали наши исследования по распределению балянусов и митилястера на биотопе, в псевдолиторали их распределение на субстрате и численность также контролируется и биотическим фактором (определяется степенью выедания рыбами балянусов).
Установлено, что выделенные типы и подтипы играют разную роль в существовании прибрежной экосистемы. В ряду: супралиторальные - псевдолиторальные - сублиторальные сообщества возрастает их роль в экосистеме (происходит переход от асообществ реципиентов к донорам).
3. Разработана типология сообществ обрастания (рис. 5). Разработанные представления о существовании разных типов сообществ обрастания прибрежья Среднего Каспия позволяют с высокой степенью вероятности прогнозировать их будущие изменения, если условия обитания изменятся.
Список опубликованных работ по теме диссертации:
- Имашова С.Н. Прибрежное сообщество обрастания в разных экологических зонах дагестанского прибрежья Среднего Каспия (монография) / С.Н. Имашова, В.А. Зайко. - Махачкала: ДГПУ, 2011.-100с.
- Имашова С.Н. Оценка влияния волнового фактора на биомассу зообентоса псевдолиторальных прибрежных сообществ обрастания, находящихся под воздействием прибоя / С.Н. Имашова, В.А. Зайко // Известия ДГПУ. Естесственные и точные науки. -а 2010. - №2. - аС. 60-63.
- Имашова С.Н. Обрастание буев в акватории махачкалинского морского порта / С.Н. Имашова, В.А. Зайко // Юг России: Экология, развитие. - 2011. - №2. - С.11-17.
- Имашова С.Н. Обрастание судов в акватории махачкалинского морского порта / С.Н.Имашова. В.А. Зайко//Известия ДГПУ. Естесственные и точные науки. - 2011. -а №2.- С. 60-63.
- Имашова С.Н. Трофические группировки макробентоса Сулакского залива Каспийского моря / С.Н. Имашова, В.А. Зайко //Экология биосистем: проблемы изучения, индикации и прогнозирования. - аАстрахань. - а2009. - С.44-45.
- Зайко В.А. Трофические группировки в обрастании махачкалинского морского порта / В.А. Зайко, С.Н. Имашова // Экология биосистем: проблемы изучения, индикации и прогнозирования. - Астрахань. - 2009. - C. 144-146.
- Имашова С.Н. Оценка времени жизни сообщества скалистой супралиторали / С.Н. Имашова // Мат. всероссийской молодежной научной конференции-школы Проблемы экологии морского шельфа, Владивосток, 16-22 сентября 2010 г. - Владивосток: Дальневосточный университет. - 2010. - С.52-54.
- Имашова С.Н. Скалистые сообщества с разным временем жизни / С.Н. Имашова, В.А. Зайко // Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий. - Краснодар: Кубанский университет. - а2010. - С. 148-149.
___________________________________________________________________
Подписано в печать 12.01.2012г.
Формат 60х841/16. Печать ризографная. Бумага офсетная.
Гарнитура Таймс. Усл. п. л. 1,5. Тираж 100 экз.
Отпечатано в издательско-типографском участке ИПЭ РД
Дахадаева 21. Тел.: 8-988-2919-920

Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]