
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
ИСКОПАЕМЫЕ ЧЕРЕПАХИ СЕМЕЙСТВ ADOCIDAE И NANHSIUNGCHELYIDAE АЗИИ: МОРФОЛОГИЯ, СИСТЕМАТИКА, ФИЛОГЕНИЯ И РАСПРОСТРАНЕНИЕ
Автореферат кандидатской диссертации
На правах рукописи
СЫРОМЯТНИКОВА
Елена Владимировна
ИСКОПАЕМЫЕ ЧЕРЕПАХИ СЕМЕЙСТВ ADOCIDAE И NANHSIUNGCHELYIDAE АЗИИ: МОРФОЛОГИЯ, СИСТЕМАТИКА, ФИЛОГЕНИЯ И РАСПРОСТРАНЕНИЕ
03.02.04 - Зоология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Санкт-Петербург
2012
Работа выполнена в Федеральном государственном бюджетном учреждении науки Зоологическом институте РАН
Научные руководители:
доктор биологических наук Суханов Владимир Борисович
кандидат биологических наук Данилов Игорь Геннадьевич
Официальные оппоненты:
Черепанов Геннадий Олегович, доктор биологических наук, доцент, Санкт-Петербургский государственный университет, кафедра зоологии позвоночных, заведующий кафедрой
Назаркин Михаил Валерьевич, кандидат биологических наук, Зоологический музей Зоологического института РАН, старший научный сотрудник
Ведущая организация:
Федеральное государственное бюджетное учреждение науки Институт проблем эволюции и экологии животных имени А.Н. Северцова РАН
Защита диссертации состоится л21 марта 2012 г. в 14 часов на заседании диссертационного совета Д 002.223.02 при Федеральном государственном бюджетном учреждении науки Зоологическом институте РАН по адресу: 199034, Санкт-Петербург, Университетская наб., д. 1, факс (812) 328-2941
С диссертацией можно ознакомиться в библиотеке Федерального государственного бюджетного учреждения науки Зоологическом институте РАН
Автореферат разослан л___ ________ 2012 г.
Ученый секретарь
диссертационного совета,
доктор биологических наук аа аа Сиделева Валентина Григорьевна
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Важные этапы эволюции скрытошейных черепах (Cryptodira) проходили в позднем мезозое - раннем кайнозое Азии, откуда известно около 13 семейств (Чхиквадзе, 1990; Yeh, 1994; Sukhanov, 2000). Среди них особое место занимают семейства Adocidae Cope, 1870 и Nanhsiungchelyidae Yeh, 1966 из мела и палеогена Азии и Северной Америки (Sukhanov, 2000; Hutchison, 2000). Несмотря на то, что известны целые скелеты адоцид и наньсюнгхелиид, их морфология остается слабо изученной, а систематическое положение ряда форм не выяснено. Особенно это касается азиатских представителей этих групп, описанных в основном по малочисленным и фрагментарным материалам. Опубликованные данные представляют собой краткие описания, зачастую без сопутствующих изображений, что затрудняет характеристику и сравнение отдельных форм. Вместе с тем многие важные материалы остаются неописанными. Систематическое положение адоцид и наньсюнгхелиид до конца не выяснено - их рассматривают в составе монофилетической группы (Adocusia), близкой к базальным эукриптодирам, или помещают среди стволовых Trionychoidea (Danilov, Parham, 2006; Joyce, 2007; Tong et al., 2009). Изучение азиатских материалов по Adocidae и Nanhsiungchelyidae имеет большое значение для решения проблем морфологии, систематики и филогении этих двух групп, а также для понимания ранних этапов эволюции скрытошейных черепах в целом.
Цель и задачи исследования.
Целью работы является изучение морфологии, филогении, распространения и построение системы черепах семейств Adocidae и Nanhsiungchelyidae Азии.
Поставленная цель определила следующие задачи:
- описание морфологии черепах семейств Adocidae и Nanhsiungchelyidae на основе новых материалов и литературных данных;
- определение положения Adocidae и Nanhsiungchelyidae в системе черепах;
- разработка филогенетической гипотезы для семейств Adocidae и Nanhsiungchelyidae;
- таксономическая ревизия азиатских Adocidae и Nanhsiungchelyidae;
- анализ геологического и географического распространения Adocidae и Nanhsiungchelyidae;
- анализ образа жизни и условий обитания Adocidae и Nanhsiungchelyidae.
Научная новизна. Проведено детальное исследование морфологии 39 видов Adocidae и Nanhsiungchelyidae (по литературным данным и коллекционным материалам). Подтверждена монофилия Adocusia и предложена схема родственных связей внутри изучаемых семейств. Ревизован и обобщен палеонтологический материал по Adocidae и Nanhsiungchelyidae из 118 местонахождений Азии в интервале поздняя юра - олигоцен. Установлен один новый род и три новых вида черепах. Уточнен состав семейств и характеристики некоторых ранее описанных таксонов; для 35 форм изменено систематическое положение. Впервые проведен подробный анализ геологического и географического распространения Adocidae и Nanhsiungchelyidae. Уточнены некоторые особенности образа жизни и условий обитания этих черепах.
Теоретическое и практическое значение работы. Результаты проведенного исследования важны для совершенствования системы скрытошейных черепах и понимания ранних этапов их эволюции. Ценность полученных данных актуальна для изучения комплексов позвоночных и древних фаун мезозоя и кайнозоя в целом. Полученные данные по распространению Adocidae и Nanhsiungchelyidae могут быть использованы для решения стратиграфических задач, реконструкций природно-климатических условий, а также для палеобиогеографии мезозоя и раннего кайнозоя Азии и Северной Америки. Результаты работы могут быть использованы в университетских курсах лекций по герпетологии, а также по зоологии и палеонтологии позвоночных.
Апробация работы. Результаты исследования были представлены на четырех Школах молодых ученых-палеонтологов (Палеонтологический институт им. А.А. Борисяка РАН (ПИН), Москва, 2006, 2007, 2008, 2009), двух Отчетных научных сессиях Зоологического института РАН (ЗИН, Санкт-Петербург, 2008, 2009), Международной конференции к 40-летию Совместной российско-монгольской палеонтологической экспедиции (СРМПЭ) (ПИН, Москва, 2009), Гэффниевском симпозиуме по черепахам (Драмхеллер, Канада, 2009), на 71-м Ежегодном съезде Общества палеонтологии позвоночных (Лас-Вегас, США, 2011), а также на научных семинарах лаборатории орнитологии и герпетологии Зоологического института РАН (2010, 2011).
Публикации. По теме диссертации опубликованы 15 работ, в том числе 5 из списка, рекомендованного ВАК.
Структура и объем диссертации. Диссертация состоит из введения, 6 глав, выводов, списка литературы и 7 приложений. Общий объем диссертации (с приложениями) - 325 страниц. Работа содержит 21 текстовый рисунок и 49 фототаблиц. Список литературы содержит 205 названий, из них 147 на иностранных языках.
Благодарности. Автор выражает глубокую признательность своим научным руководителям кандидату биологических наук И.Г. Данилову (ЗИН РАН) и доктору биологических наук В.Б. Суханову (ПИН РАН, Москва) за предоставление материала и неоценимую помощь в работе. Я благодарю следующих коллег за всестороннюю помощь и участие: А.О. Аверьянова (ЗИН РАН), М. Борсук-Бялыницку (Варшава, Польша), А.Р. Соколова, Н.М. Кадлец и Т.В. Куражеву (Санкт-Петербург), Е.Н. Мащенко (ПИН РАН), Д. Бринкмана и Б. Стрилиски (Драмхеллер, Канада), Р. Хираяму (Токио, Япония), Х. Тонг и Ф. де Лаппарент де Бруан (Париж, Франция), У. Джойса (Тюбинген, Германия), Т. Шейера (Цюрих, Швейцария), П. Холройд и Х. Хатчисона (Беркли, США), О. Матеуса (Лориньян, Португалия), В.В. Титова (Ростов-на-Дону, Россия), а также участников международной палеонтологической экспедиции URBAC (1997Ц2006). Я признательна коллективам лаборатории териологии и отделения герпетологии лаборатории орнитологии и герпетологии ЗИН РАН за поддержку и создание прекрасных условий для выполнения настоящей работы.
СОДЕРЖАНИЕ РАБОТЫ
ГЛАВА 1. ИСТОРИЯ ИЗУЧЕНИЯ ADOCIDAE И NANHSIUNGCHELYIDAE
Изучение черепах семейств Adocidae и Nanhsiungchelyidae началось более 150 лет назад с описания первых находокAdocusиз позднего мела Северной Америки (Leidy, 1856, 1865). Для них было выделено отдельное семейство Adocidae, куда впоследствии были отнесены также другие близкие к Adocus североамериканские роды (Cope, 1870, 1871, 1875). Семейство Nanhsiungchelyidae было установлено значительно позднее и первое время включало только единственный азиатский род Nanhsiungchelys (Yeh, 1966). Только более десяти лет спустя к этому семейству были отнесены и другие таксоны, многие из которых ранее рассматривались в составе адоцид (Несов, 1977; Несов, Юлинен, 1977 и др.). При этом состав обоих семейств неоднократно менялся и пополнялся новыми формами, а сами семейства даже на время выходили из употребления (Нау, 1908, 1910; Gilmore, 1919; White, 1972). В 70, 80-е годы прошлого века в литературе стали появляться многочисленные упоминания находок адоцид и наньсюнгхелиид из Азии, включающие описания их новых таксонов (Хозацкий, Несов, 1977; Несов, 1981, 1997; Несов, Красовская, 1984 и др.). При этом многие из них устанавливались по фрагментарным и малочисленным материалам. С началом использования кладистического метода в изучении черепах появляются работы по определению родственных связей Adocidae и Nanhsiungchelyidae. Первые филогенетические исследования (Gaffney, Meylan, 1988; Meylan, Gaffney, 1989) показали положение Adocidae в качестве сестринской группы к кладе, объединяющей Nanhsiungchelys (Nanhsiungchelyidae) и Trionychia (Trionychidae + Carettochelyidae). Дальнейшие работы в большинстве своем были посвящены филогении Nanhsiungchelyidae, постепенно включая новые таксоны этого семейства в кладистические исследования (Brinkman, Nicholls, 1993; Brinkman, Peng, 1996; Brinkman, 1998; Hirayama et al., 2001; Joyce, Norell, 2005). Наряду с этим, филогения Adocidae в литературе разработана очень слабо (Lapparent de Broin, 2004). Недавние работы показали существование клады Adocusia (Adocidae + Nanhsiungchelyidae +Yehguia), занимающей сестринское положение к Trionychia (Danilov, Parham, 2006, 2008; Joyce, 2007; Sterli, 2010). Однако в одном из последних таких исследований монофилия Adocusia не подтверждается (Tong et al., 2009).
Приведенный в данной главе обзор показывает, что, несмотря на достаточно долгий период изучения адоцид и наньсюнгхелиид и многочисленность их находок, сведения по морфологии, систематике и филогении этих групп представлены очень неполно, разрозненны и зачастую противоречивы.
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ
2.1. Материал. Использованный в работе материал по Adocidae и Nanhsiungchelyidae собирался автором в нескольких местонахождениях Центральных Кызылкумов (Узбекистан, 2003 г.) и изучался в коллекциях ряда учреждений (ЗИН РАН, Санкт-Петербург; ПИН РАН, Москва; Центральный научно-исследовательский геолого-разведочный музей им. Ф.Н. Чернышева, Санкт-Петербург; Институт палеобиологии Польской АН, Варшава, Польша; Национальный музей естественной истории, Париж, Франция; Королевский музей Тиррела, Драмхеллер, Канада; Музей палеонтологии Калифорнийского университета, Беркли, США). Материал включал около 10000 панцирных и других элементов скелета. Из них около 9000 принадлежали семейству Adocidae, остальные - Nanhsiungchelyidae. Большая часть материалов была представлена изолированными костными пластинками или реже более крупными фрагментами панциря от различных особей черепах. Азиатские материалы происходят из 118 известных на данный момент местонахождений с территорий Казахстана, Киргизии, Китая, Лаоса, Монголии, Таджикистана, Тайланда, Туркменистана, Узбекистана и Японии. На основе коллекционных материалов были изучены 18 (13 видов Adocidae и 5 видов Nanhsiungchelyidae) из 39 видов, а также многочисленные адоциды и наньсюнгхелииды неопределенного систематического положения. Остальные виды рассматриваемых семейств изучались по литературным данным и оригинальным изображениям.
2.2. Методы. Материалы частично отбирались из неразобранных сборов, при необходимости очищались от породы и склеивались. Определение материалов по Adocidae и Nanhsiungchelyidae осуществлялось по всему набору известных признаков. Схема описания, а также номенклатура черепных элементов дается по Дж. Гэффни (Gaffney, 1979) и В.Б. Суханову (1964), костных и роговых элементов панциря - по Л.И. Хозацкому и В.Б. Суханову (1973) с изменениями по Х. Хатчисону и Д. Брамблу (Hutchison, Bramble, 1981). Измерения плечевых костей проводились по методике Р. Зангерла (Zangerl, 1953).
Анализ родственных связей внутри Adocidae и Nanhsiungchelyidae и уточнение положения семейств в системе черепах проводился с применением кладистического анализа. Таксон-признаковая матрица составлялась в программе NDE 0.5.0 (Page, 2001) и анализировалась программой PAUP 4.0b10 (Swofford, 2002). Для уточнения положения Adocidae и Nanhsiungchelyidae в системе черепах использовался алгоритм Heuristic Search, для анализа взаимоотношений внутри Adocidae и Nanhsiungchelyidae - алгоритм Branch-and-Bound. Всем признакам придавался равный вес, признаки не ордировались и для всех них допускалась реверсия. Индексы Бремер-поддержки (Bremer supports) рассчитывались с использованием программы Autodecay 4.0.1 (Eriksson, 1998).
ГЛАВА 3. МОРФОЛОГИЯ ADOCIDAE И NANHSIUNGCHELYIDAE
В данной главе на основе коллекционных материалов и литературных данных подробно описывается морфология Adocidae и Nanhsiungchelyidae, а также проводится отбор признаков для разработки филогенетической гипотезы для обоих семейств (см. Главу 4).
3.1. Череп. По пропорциям череп Adocidae и Nanhsiungchelyidae в целом сходен с Dermatemydidae и сочетает в себе как примитивные (небные кости не контактируют по средней линии, каналы для мозговой и небной ветвей внутренней сонной артерии примерно равны по диаметру), так и прогрессивные (трохлеарный отросток хорошо развит, имеются верхнечелюстные зубы (maxillary tooth), foramen posterius canalis carotici interni полностью сформирован крыловидными костями) признаки (Meylan, Gaffney, 1989; Brinkman, 1998). Внутри рассматриваемых семейств была выявлена изменчивость по форме черепа, размерам трохлеарного отростка, щечной вырезки, наличию/отсутствию участия лобных костей в формировании глазницы, форме верхнечелюстных костей, форме и контактам элементов неба. У всех известных адоцид и наньсюнгхелиид нижняя челюсть характеризуется наличием нижнечелюстного кармана (dentary pocket). При этом зубная кость у большинства из них имеет узкую альвеолярную поверхность, значительно суженную спереди и слегка расширенную сзади, тогда как у Basilemys ее ширина примерно одинакова по всей длине. Высокий и хорошо развитый венечный отросток, по-видимому, характерен только для Adocus, тогда как у Shachemys наблюдается относительно низкий венечный отросток, смещенный в заднюю половину ветви нижней челюсти.
3.2. Панцирь. Панцирь Adocidae и Nanhsiungchelyidae, как и у большинства черепах, представлен хорошо развитыми карапаксом и пластроном, покрытыми скульптурой из мелких (меловые Adocus, Ferganemys) или крупных (Nanhsiungchelyidae) ямок (Sukhanov, 2000). Для обоих семейств были выявлены различия по ширине пигальной пластинки (узкая у Adocidae и широкая у Nanhsiungchelyidae), длине мостов пластрона (короткие у Adocidae и длинные у большинства Nanhsiungchelyidae), длине передней доли пластрона (короткая у Adocidae и длинная у Nanhsiungchelyidae) и ряду других признаков. Установлено, что строение панциря примитивных Adocidae (Yehguia) характеризуется нормально развитыми головками ребер и реберными валиками и захождением маргинальных щитков на реберные пластинки в средней части карапакса. У более продвинутых форм происходит усиление захождения щитков, продлеваемое на заднюю часть карапакса (Adocinae) или исчезновение этого захождения (Shachemydinae). Строение панциря примитивных Nanhsiungchelyidae (Kharakhutulia) характеризуется слабой нухальной вырезкой, сокращенной невральной формулой, широким I центральным щитком, низким положением плевро-маргинальной борозды, короткой передней долей пластрона и слабым захождением гулярных щитков на дорзальную сторону пластрона. У более продвинутых форм наблюдается восстановление невральной формулы, сужение I центрального щитка и удлинение передней доли пластрона. Скульптура в виде точек, представляющих собой отверстия каналов, пронизывающих костные пластинки и известная только у Shachemys, была выявлена у азиатских и североамериканских палеогеновых Adocus. Внутри рассматриваемых семейств наблюдается значительная изменчивость по форме и количеству роговых (захождению маргинальных щитков на реберные пластинки, форме и размеру прецентрального и пластральных щитков) и костных (количеству невральных пластинок, степени отгиба свободного края краевых пластинок) элементов. Наличие ряда признаков в строении Adocidae и Nanhsiungchelyidae (редукция серии невральных пластинок, увеличенные в высоту задние краевые пластинки, захождение маргинальных щитков на реберные пластинки в средней части панциря) позволяют сближать их с примитивными эукриптодирами уровня Xinjiangchelyidae. Помимо этого, Adocidae и Nanhsiungchelyidae сходны с Xinjiangchelyidae по признакам (невральная формула 6>4<6<6<6<6<6, извилистая срединная борозда пластрона), которые прежде считались синапоморфиями для Adocusia (Joyce, 2007).
3.3. Непанцирный посткраний. Строение непанцирного посткрания Adocidae и Nanhsiungchelyidae в целом характерно для других черепах (Суханов, Нармандах, 1977; Meylan, Gaffney, 1989; Brinkman, Peng, 1996; Brinkman, 2003; Joyce, Norell, 2005). Среди основных особенностей выделяется наличие опистоцельных шейных позвонков, известных еще только у Trionychoidea (Meylan, 1987). В строении костей поясов и свободных конечностей изучаемых семейств можно отметить следующий ряд различий: кости относительно тонкие и удлиненные (Adocidae) или более короткие и широкие (Nanhsiungchelyidae); акромиальный отросток прямой (Adocidae) или скрученный в средней части (Nanhsiungchelyidae); коракоид длинный узкий (Adocidae) или широкий дистально (Nanhsiungchelyidae); симфиз лобковых костей длинный (Adocidae) или короткий (Nanhsiungchelyidae); фаланговая формула 2-3-3-3-3, фаланги пальцев удлиненные (Adocidae) или 1-2-2-2-2, фаланги укороченные (Nanhsiungchelyidae); конечности покрыты остеодермами (Nanhsiungchelyidae) или остеодермы отсутствуют (Adocidae).
ГЛАВА 4. ФИЛОГЕНИЯ ADOCIDAE И NANHSIUNGCHELYIDAE
4.1. Положение Adocidae и Nanhsiungchelyidae в системе черепах.
Для уточнения положения Adocidae и Nanhsiungchelyidae в системе черепах были проведены два кладистических анализа на основе данных из нескольких работ (Danilov, Parham, 2006; Joyce, 2007; Mateus et al., 2009; Tong et al., 2009).
Для Анализа 1 была использована матрица из трех предшествующих работ (Danilov, Parham, 2006; Joyce, 2007; Mateus et al., 2009). В эту матрицу, включающую 143 признака для 71 таксона были внесены следующие изменения: добавлены один таксон Adocidae (Ferganemysverzilini),два таксона Nanhsiungchelyidae (KharakhutuliakalandadzeiиNanhsiungchelyswuchingensis) и таксон неясного систематического положения - Basilochelys macrobios Tong et al., 2009;добавлен один признак - скульптура панциря в виде ямок, разделенных гребнями: (0) нет, (1) есть; для некоторых таксонов (Adocusbeatusи Zangerlianeimongolensis) изменены кодировки отдельных признаков. Итоговая матрица включала 144 признака для 75 таксонов. В результате филогенетического анализа получено 374 дерева, строгий консенсус этих деревьев представлен на рис. 1. Данный анализ показывает, что представители Adocidae и Nanhsiungchelyidae образуют кладу Adocusia, которая поддерживается тремя признаками: наличие невральной формулы 6>4<6<6<6<6<6, сильная извилистость срединной борозды пластрона, скульптура панциря в виде ямок, разделенных гребнями. В отличие данных из предыдущих работ, первые два из перечисленных признаков более не являются синапоморфиями Adocusia (см. Главу 3). Отношения внутри полученной клады не разрешены, однако данный анализ не направлен на решение этого вопроса.
В составе Adocusia оказалсяBasilochelysmacrobiosиз поздней юры - раннего мела Тайланда (Tong et al., 2009), что вызвано сочетанием трех признаков: контакт между сошником и крыловидными костями отсутствует, небные кости посередине контактируют друг с другом; невральная формула 6>4<6<6<6<6<6. Последний из этих признаков не уникален для Adocusia (Главу 3). Наряду с этим, Basilochelysдемонстрирует ряд признаков (амфицельные шейные позвонки, длинное первое ребро, парные ямки на вентральной поверхности базисфеноида), наблюдаемых у базальных эукриптодир. Таким образом, положение Basilochelys остается спорным (см. Tong et al., 2009) и его включение в состав Adocusia не имеет достаточных оснований. По-видимому, Basilochelys следует рассматривать в составе базальных эукриптодир, развивающихся от уровня Xinjiangchelyidae в направлении Adocidae и Nanhsiungchelyidae.

Рис. 1. Консенсусное дерево (длина шагов = 142, CI = 0.66, RI = 0.88), полученное в результате Aнализа 1. Показаны только Eucryptodira.
Клада Adocusia занимает сестринское положение по отношению к Peltochelys + Trionychoidea, что поддерживается наличием следующих признаков: экстрагулярные щитки имеются (не применимо для Trionychoidea); сочленения между IVЦVII позвонками опистоцельные. Наличие экстрагулярных щитков следует рассматривать как примитивную черту, а опистоцельные шейные позвонки могли возникать независимо в разных группах черепах. Поэтому сближение Adocusia и Trionychoidea требует дополнительных обоснований. Топология остальных клад отличается от таковой в предыдущих работах, однако в данной работе мы это не обсуждаем.
Для Анализа 2 была использована матрица из работы Х. Тонг с соавторами (Tong et al., 2009). Эта матрица, включающая 74 признака для 25 таксонов, была подвергнута следующим изменениям: добавлены два таксона Adocidae (Yehguia, Ferganemys) и один Nanhsiungchelyidae (Kharakhutulia); изменена формулировка признака скульптура панциря: добавлено состояние 2 (скульптура в виде ямок, разделенных гребнями); для некоторых таксонов (Adocus, Basilemys,Zangerliaи Xinjiangchelys) изменены кодировки отдельных признаков. Итоговая матрица включала 75 признаков для 28 таксонов. В результате филогенетического анализа получено 4 дерева, строгий консенсус этих деревьев представлен на рис. 2.

Рис. 2. Консенсусное дерево (длина шагов = 187, CI = 0.44, RI = 0.73), полученное в результате Aнализа 2.
В отличие от анализа, проведенного Х. Тонг с соавторами, в результате нашего анализа Adocidae (в составе Yehguia, Adocus и Ferganemys) и Nanhsiungchelyidae (в составе Basilemys, Kharakhutulia, NanhsiungchelysиZangerlia) образовали кладу Adocusia. В составе Adocidae отношения между таксонами не разрешены, тогда как представители Nanhsiungchelyidae образовали кладу (Kharakhutulia + (Basilemys + (Nanhsiungchelys + Zangerlia))). Adocusia поддерживается единственной синапоморфией - наличием скульптуры панциря в виде ямок, разделенных гребнями. Adocidae объединяются по наличию значительного участия теменных костей в образовании трохлеарного отростка, а Nanhsiungchelyidae - расширенного вентромедиального края VI маргинального щитка. Клада (Basilemys + (Nanhsiungchelys + Zangerlia)) поддерживается двумя синапоморфиями: широкое захождение гулярных щитков на утолщенную переднюю пластральную губу; передняя доля пластрона выступает вперед за край карапакса.
Клада Adocusia группируется с Basilochelys по наличию невральной формулы 6>4<6<6<6<6<6, однако положение Basilochelys является спорным (см. выше). Клада Adocusia + Basilochelys занимает сестринское положение по отношению к Trionychoidea (Trionychidae + Carettochelyidae) по наличию ретроартикулярного отростка (неизвестно для Basilochelys), опистоцельного или двояковыпуклого восьмого шейного позвонка (неизвестно для Basilochelys), ямчатой, гребнистой или червеобразной скульптуры панциря. Вопрос сближения Adocusia и Trionychoidea обсуждается выше.
Проведенные филогенетические исследования положения Adocidae и Nanhsiungchelyidae в системе черепах (Анализы 1 и 2) подтверждают существование клады (монофилетической группы) Adocusia, которая в ранговой систематике соответствует надсемейству Adocoidea.
4.2. Филогенетические отношения в семействах Adocidae и Nanhsiungchelyidae.
Анализ 3. Филогенетическая гипотеза для Adocidae и Nanhsiungchelyidae разрабатывалась на основе данных из работы У. Джойса и М. Норелла (Joyce, Norell, 2005) с дополнениями из других публикаций (Meylan, Gaffney, 1989; Brinkman, Peng, 1996; Brinkman, 1998; Hirayama et al., 2001; Joyce, Norell, 2005; Tong, Mo, 2010). Итоговая матрица включала 75 остеологических признаков для 26 таксонов. В результате филогенетического анализа были получены 11 деревьев, строгий консенсус этих деревьев представлен на рис. 3. Результаты анализа подтверждают монофилию Adocidae, Nanhsiungchelyidae, Adocinae и Shachemydinae, которые образуют единую кладу Adocusia (= Adocoidea). Монофилия Adocusia поддерживается наличием шовного соединения пластрона с карапаксом. Признаки невральная формула 6>4<6<6<6<6<6 и сильная извилистость срединной борозды пластрона больше не входят в число синапоморфий Adocusia (см. Главу 3). Ранее монофилия Adocusia основывалась на данных только по 2Ц4 представителям этой группы, тогда как в нашем анализе учтены данные по большинству Adocidae и Nanhsiungchelyidae. В результате проведенного анализа Yehguia tatsuensis включается в состав семейства Adocidae в качестве наиболее примитивного представителя. Дальнейшее разделение Adocidae на две клады соответствует принятому делению семейства на подсемейства Adocinae и Shachemydinae. Подсемейство Adocinae включает один род Adocus, его азиатские представители образуют кладу Adocusaksary + A. amtgai. Выделение A. amtgaiв самостоятельный родAdocoides (Sukhanov, Narmandakh, 2006) не поддерживается. Подсемейство Shachemydinae представлено родами Ferganemys, Shachemys и видом неопределенной родовой принадлежности - УFerganemysФ itemirensis, ранее принятая монофилия рода Ferganemys не подтверждается. В составе родаShachemys

Рис. 3. Консенсусное дерево (длина шагов = 67, CI = 0.81, RI = 0.92), полученное в результате Aнализа 3. Номера обозначают индексы Бремер-поддержки.
выделяется клада Sh. ancestralis + Sh. baibolatica. В составе Nanhsiungchelyidae наиболее базальное положение занимает Kharakhutuliakalandadzei,остальные таксоны образуют клады Anomalochelys + Nanhsiungchelys, Basilemys, Hanbogdemys + Jiangxichelys, УZangerliaФ neimongolensis + УZ.Ф dzamynchondi, образующими политомию с Z. testudinimorpha и УZ.Ф ukhaachelys (выделяемая ранее клада Zangerlia не поддерживается). Результаты проведенного анализа опровергли утверждение о существовании монофилетичной азиатской клады Nanhsiungchelyidae (Joyce, Norell, 2005). Очевидно, что азиатские представители семейства дали начало североамериканским сородичам. Однако предложенная гипотеза не позволяет установить родственные связи Basilemys с какой-либо из азиатских групп Nanhsiungchelyidae.
ГЛАВА 5. СИСТЕМАТИКА НАДСЕМЕЙСТВА ADOCOIDEA
В данной главе приводятся характеристики таксонов надсемейства Adocoidea (Adocidae + Nanhsiungchelyidae) известных в настоящее время с территории Азии. В составе надсемейства рассматриваются 2 семейства, 11 родов, 26 видов, из которых 1 род и 3 вида новые, для 35 форм изменено систематическое положение, описаны 20 новых находок.
Отряд Testudines Batsch, 1788
Подотряд Cryptodira Cope, 1868
Надсемейство Adocoidea Cope, 1870
Краткая характеристика: скульптура панциря в виде ямок, разделенных гребнями. Состав: семейства Adocidae Cope, 1870 и Nanhsiungchelyidae Yeh, 1966. Распространение: поздняя юра - олигоцен Азии, поздний мел - палеоцен Северной Америки.
Семейство Adocidae Cope, 1870
Краткая характеристика: скульптура панциря в виде маленьких ямок с низкими гребнями; пигальная пластинка длинная и узкая; головки ребер слабо развиты; реберные валики не выражены; роговые борозды узкие и неглубокие. Состав: подсемейства Adocinae Cope, 1870 и Shachemydinae Khosatzky in Nessov et Khosatzky, 1977; роды неопределенного систематического положения: Isanemys Tong et al., 2006; Yehguia Danilov et Parham, 2006. Распространение: поздняя юра - олигоцен Азии, поздний мел - палеоцен Северной Америки.
Подсемейство Adocinae Cope, 1870
Краткая характеристика: свободный край краевых пластинок с отгибом; маргинальные щитки заходят на реберные пластинки начиная с IIIЦV пары. Состав: Adocus Cope, 1868 (= AdocoidesSukhanov et Narmandakh, 2006; = Shineusemys Sukhanov et Narmandakh, 2006). Распространение: баррем - кампан Азии, коньяк - палеоцен Северной Америки.
Род Adocus Cope, 1868
Краткая характеристика: см. характеристику подсемейства. Состав: A. agilis Cope, 1868; A. aksaryNessov in Nessov et Krasovskaya, 1984; A. amtgaiNarmandakh, 1985; A. annexus (Hay, 1910); A. beatus (Leidy, 1865); A. bossi Gilmore, 1919; A. bostobensisSyromyatnikova et Danilov,2009; A. dzhurtasensisSyromyatnikova et Danilov,2009; A. foveatusNessov et Khosatzky in Khosatzky et Nessov, 1977; A. hesperius Gilmore, 1919; A. kirtlandius Gilmore, 1919; A. kizylkumensisNessov, 1981; A. microglypha (Cope, 1871); A. onerosus Gilmore, 1919; A. planus (Sukhanov et Narmandakh, 2006); A. striatula (Cope, 1870); A. substrictus (Hay, 1908). Распространение: см. распространение подсемейства.
AdocusaksaryNessov in Nessov et Krasovskaya, 1984
Краткая характеристика: V centrale относительно узкое; маргинальные щитки заходят на реберные пластинки с III или IV marginalia; медиальные края экстрагулярных щитков длинные. Распространение: поздний турон Узбекистана.
AdocusamtgaiNarmandakh, 1985
Краткая характеристика: IIЦIV pleuralia короткие (их длина составляет около 30% ширины); маргинальные щитки заходят на реберные пластинки с IV marginalia. Распространение: поздний турон - сантон Монголии. Замечания: A. amtgai был выделен в самостоятельный род Adocoides Sukhanov et Narmandakh, 2006 (Sukhanov, 2000; Sukhanov, Narmandakh, 2006), однако по результатам проведенного филогенетического анализа этот вид рассматривается в составе рода Adocus (Syromyatnikova et al., 2011).
Adocus bostobensis Syromyatnikova et Danilov, 2009
Краткая характеристика: отличается от других видов рода крупными размерами панциря, а от азиатских видов - формой прецентрального щитка. Распространение: сантон - ранний кампан Казахстана.
Adocus dzhurtasensis Syromyatnikova et Danilov, 2009
Краткая характеристика: нухальная вырезка сильно развита; передние края загривковой пластинки и прецентрального щитка узкие. Распространение: сантон - ранний кампан Казахстана.
Adocusfoveatus Nessov et Khosatzky in Khosatzky et Nessov, 1977
Краткая характеристика: передний край эпипластронов широкий; маргинальные щитки заходят на реберные пластинки с V marginalia; gularia расширены спереди; extragularia укорочены медиально. Распространение: сантон - ранний кампан Казахстана.
AdocuskizylkumensisNessov, 1981
Краткая характеристика: нухальная вырезка отсутствует; прецентральный щиток трапециевидный, расширенный сзади. Распространение: ранний сеноман Узбекистана.
Adocus planus (Sukhanov et Narmandakh, 2006)
Краткая характеристика: абдоминальные щитки меньше по длине по сравнению с феморальными. Распространение: сеноман - ранний турон Монголии. Замечания: первоначально этот вид был установлен в составе самостоятельного рода Shineusemys Sukhanov et Narmandakh, 2006 (Sukhanov, 2000; Sukhanov, Narmandakh, 2006). Однако указанные для этого рода диагностичные признаки значительно варьируют среди адоцид. По всем другим особенностям рассматриваемый вид вполне укладывается в рамки Adocus sensu lato.
Adocus sp.
К Adocus sp. 1Ц9 отнесены фрагментарные находки Adocusиз мела Монголии, Узбекистана и Японии, не позволяющие определить их принадлежность к какому-либо из известных видов.
Подсемейство Shachemydinae Khosatzky in Nessov et Khosatzky, 1977
Краткая характеристика: небные кости контактируют по средней линии; маргинальные щитки не заходят на реберные пластинки; энтопластрон укорочен; экстрагулярно-гумеральная борозда приближена к эпи-энтогиопластральному шву или совпадает с ним. Состав: Ferganemys Nessov et Khosatzky, 1977; ShachemysKuznetsov, 1976. Распространение: мел Азии.
РодFerganemys Nessov et Khosatzky, 1977
Краткая характеристика: прецентральный щиток широкий и короткий; задняя доля пластрона относительно длинная и сужающаяся назад; инфрамаргинальных щитков одна или две пары. Состав: Ferganemysverzilini Nessov et Khosatzky, 1977. Распространение: альб Киргизии. Замечания: В составе этого рода мы рассматриваем только типовой вид Ferganemysverzilini. По результатам филогенетического анализа F. itemirensisобразует политомию с F. verzilini и Shachemys и поэтому рассматривается как УFФ. itemirensis (см. ниже).
Ferganemysverzilini Nessov et Khosatzky, 1977
Краткая характеристика: см. характеристику рода. Распространение: см. распространение рода.
РодShachemysKuznetsov, 1976
Краткая характеристика: невральные пластинки отсутствуют или представлены только первым элементом; прецентральный щиток отсутствует; эпи-энтогиопластральный шарнир имеется; передний край энтопластрона прямой; скульптура панциря точечная. Состав:Sh. ancestralis Nessov in Nessov et Krasovskaya, 1984; Sh. baibolaticaKuznetsov, 1976; Sh. laosiana Lapparent de Broin, 2004. Распространение: конец раннего мела Лаоса и Тайланда, поздний мел Казахстана, Средней Азии и Японии.
Shachemys ancestralis Nessov in Nessov et Krasovskaya, 1984
Краткая характеристика: загривковая пластинка относительно короткая; передняя доля пластрона слабо сужается спереди. Распространение: поздний турон Узбекистана. Замечания: этот вид первоначально был установлен как подвид Sh. baibolaticaancestralis (Несов, Красовская, 1984), однако новые данные по морфологии этой формы позволяют повысить ее ранг до видового.
ShachemysbaibolaticaKuznetsov, 1976
Краткая характеристика: загривковая пластинка относительно длинная; передняя доля пластрона сильно сужается спереди. Распространение: ранний сантон Таджикистана, сантон - ранний кампан Казахстана.
ShachemyslaosianaLapparrent de Broin, 2004
Краткая характеристика: нухальная вырезка отсутствует; IIЦIV центральные щитки шире плевральных; маргинальные щитки относительно высокие. Распространение: апт - альб Лаоса. Замечания: материал по aff. Xinjiangchelyssp. (Lapparrent de Broin, 2004) относится к Shachemys на наличии точечной скульптуры панциря, форме эпипластронов и энтопластрона. Однако данная форма имеет ряд отличий от Sh. laosianaи, возможно, принадлежит к отдельному виду.
Shachemys sp.
К Shachemys sp. 1Ц8 отнесены фрагментарные находки Shachemys из мела Казахстана, Таджикистана, Тайланда, Узбекистана и Японии,не позволяющие определить их принадлежность к какому-либо из известных видов.
Shachemydinae inc. gen.
"Ferganemys" itemirensis Nessov, 1981
Краткая характеристика: I neurale четырехугольное, нерасширенное сзади; свободные края загривковой и краевых пластинок без отгиба; прецентральный щиток длинный и трапециевидный или отсутствует; инфрамаргинальных щитков четыре пары. Распространение: ранний сеноман Узбекистана. Замечания: см. Замечания к роду Ferganemys.
Shachemydinae indet.
К Shachemydinae indet. 1 отнесен эпипластрон и фрагменты реберных пластинок, описанные ранее как отдельный род и вид MlynarskiellamarianiShuvalov et Chkhikvadze, 1986 (Шувалов, Чхиквадзе, 1986). По своей морфологии (пропорциям, расположению роговых борозд и скульптуре поверхности) эпипластрон сходен с таковыми у представителей Shachemydinae. Слияние гулярных щитков, указанное в качестве диагностического признака этого таксона, не может быть достоверно установлено по единственному элементу. Это не позволяет определять материал точнее, чем Shachemydinae indet, а M. mariani рассматривается как nomen dubium. К Shachemydinae indet. 2 отнесен материал из нижнего мела (валанжин - готерив) Японии, описанный как Trionychoids A and B (Hirayama, 2010). Принадлежность к Shachemydinae основана на наличии ямчатой скульптуры панциря, длинной пигальной пластинки, пропорциях эпипластронов и энтопластрона, слабых и узких роговых бороздах.
Adocidae inc. subfam. et gen.
УAdocusФ kazachstanica Chkhikvadze, 1973
Краткая характеристика: нухальная вырезка слабо развита; передний край загривковой пластинки относительно широкий; прецентральный щиток очень узкий; скульптура панциря в виде точек и др. Распространение: средний эоцен Казахстана. Замечания: УAdocusФkazachstanica демонстрирует наличие точечной скульптуры панциря, отличной от ямчатой скульптуры меловых Adocus.
УAdocusФ orientalis Gilmore, 1931
Краткая характеристика: передний край пластрона прямой; ширина переднего края эпипластрона больше длины эпипластрального симфиза; экстрагулярные щитки короткие; скульптура панциря в виде точек. Распространение: поздний эоцен Монголии и Китая. Замечания:По этому виду были описаны новые материалы из позднего эоцена Монголии. УAdocusФorientalis демонстрирует наличие точечной скульптуры панциря, отличной от ямчатой скульптуры меловых Adocus.
cf. УAdocusФ orientalis
Замечания: отнесение cf. УAdocusФ orientalis к УAdocusФ основано на форме передней части гиопластрона и скульптуре панциря в виде точек, характерных для палеогеновых адокусов. Эта форма сходна с УA.Ф orientalisпо пропорциям и щиткованию пластрона, однако пока мы воздерживаемся от отнесения ее к УA.Ф orientalis.
УAdocusФ sp.
К УAdocusФ sp. 1Ц3 отнесены фрагментарные находки УAdocusФ из палеоцена и олигоцена Казахстана и Китая,не позволяющие определить их принадлежность к какому-либо из известных видов.
Adocidae indet.
К Adocidae indet. 1Ц3 отнесены фрагментарные находки Adocidae из верхнего мела Узбекистана и нижнего мела Японии, не позволяющие определить их принадлежность к какому-либо из известных родов.
Семейство Nanhsiungchelyidae Yeh, 1966
Краткая характеристика: скульптура панциря в виде крупных ямок с высокими гребнями; пигальная пластинка короткая; пекторальные щитки не участвуют в образовании подмышечных вырезок; роговые борозды широкие и глубокие. Состав:роды AnomalochelysHirayama et al., 2001; Basilemys Hay, 1902; HanbogdemysSukhanov et Narmandakh, 2006; Jiangxichelys Tong et Mo, 2010; Kharakhutulia Sukhanov et al., 2008;Nanhsiungchelys Yeh, 1966;Zangerlia Mlynarski, 1972. Распространение: мел Азии и Северной Америки.
Род Hanbogdemys Sukhanov et Narmandakh, 2006
Краткая характеристика: нухальная вырезка хорошо развита, образована загривковой и I краевыми пластинками; I centrale не расширено спереди и не контактирует со II marginalia; передняя доля пластрона клиновидная. Состав:H. jaganchobili(Sukhanov et Narmandakh 2006) иH. orientalisSukhanov et Narmandakh, 1975. Распространение: сеноман - кампан Монголии.
Hanbogdemys jaganchobili (Sukhanov et Narmandakh, 2006)
Краткая характеристика: карапакс сужен сзади; нухальная вырезка относительно широкая; прецентральный щиток относительно небольшой. Распространение: кампан Монголии. Замечания: Первоначально этот вид был установлен в составе рода BulganemysSukhanov et Narmandakh, 2006 (Sukhanov, 2000; Sukhanov, Narmandakh, 2006), однако по ряду признаков мы относим его к роду Hanbogdemys.
Hanbogdemyssp.
К Hanbogdemyssp. 1Ц4 отнесены фрагментарные находки Hanbogdemys из верхнего мела Монголии, не позволяющие определить их принадлежность к какому-либо из известных видов. Из них находки Hanbogdemys в сеномане указываются впервые.
Род KharakhutuliaSukhanov, Danilov et Syromyatnikova, 2008
Краткая характеристика: нухальная вырезка слабо развита; невральных пластинок семь; I centrale расширено спереди; передняя доля пластрона короткая; гулярная и анальная вырезки имеются. Состав: KharakhutuliakalandadzeiSukhanov et al, 2008. Распространение: сеноман - ранний турон Монголии.
KharakhutuliakalandadzeiSukhanov, Danilov et Syromyatnikova, 2008
Краткая характеристика: см. характеристику рода. Распространение: см. распространение рода.
РодZangerlia Mlynarski, 1972
Краткая характеристика: бугор в задней части карапакса имеется; расстояние от конца задней доли пластрона до заднего края карапакса больше длины задней доли пластрона; инфрамаргинальные щитки не образуют непрерывный ряд. Состав: ZangerliatestudinimorphaMlynarski, 1972. Распространение: кампан - маастрихт Монголии. Замечания: в составе этого рода мы рассматриваем только типовой вид Z. testudinimorpha, а другие три вида, ранее относимые к этому роду, по результатам филогенетического анализа образуют политомию с другими родами Nanhsiungchelyidae и рассматриваются как УZangerliaФ.
ZangerliatestudinimorphaMlynarski, 1972
Краткая характеристика: см. характеристику рода. Распространение: см. распространение рода. Замечания: Z. testudinimorpha была описана М. Млынарским (Mlynarski, 1972) по неполному панцирю из местонахождения Нэмэгт. В той же работе к этому виду был причислен неполный карапакс из одновозрастного местонахождения Хулсан, который рассматривается нами как Nanhsiungchelyidae indet.
Nanhsiungchelyidae inc. gen.
УZangerliaФ dzamynchondi Sukhanov et Narmandakh, 2006
Краткая характеристика: I супрапигальная пластинка широкая; плевро-маргинальная борозда отдалена от косто-периферального шва. Распространение: кампан Монголии. Замечания: см. выше.
Nanhsiungchelyidae indet.
К Nanhsiungchelyidae indet. 1Ц19 отнесены фрагментарные находки Nanhsiungchelyidae из мела Китая, Монголии, Узбекистана и Японии, не позволяющие определить их принадлежность к какому-либо из известных родов. К Nanhsiungchelyidae indet. 6 отнесен материал из верхнего мела (сеноман - нижний турон) Монголии, выделенный в отдельный род и вид Charitonyxtajanikolaevae Ckhikvadze, 1980 (Чхиквадзе, Шувалов, 1980; Ckhikvadze, 1987). Ch. tajanikolaevae в целомсходен с азиатскими Nanhsiungchelyidae, однако ряд признаков отличает его от остальных Nanhsiungchelyidae. Более того, часть материалов проявляет сходство с Trionychidae (Chkhikvadze, 1980, p. 502), и, вероятно, не принадлежат Ch. tajanikolaevae.Фрагментарный материал по этому видуне позволяет определить его точнее, чем Nanhsiungchelyidae indet.,аCh. tajanikolaevae рассматривается как nomen dubium.
ГЛАВА 6. РАСПРОСТРАНЕНИЕ, УСЛОВИЯ ОБИТАНИЯ И ОБРАЗ ЖИЗНИ ADOCIDAE И NANHSIUNGCHELYIDAE
6.1. Геологическое и географическое распространение. В Азии Adocidae и Nanhsiungchelyidae отличаются довольно широким распространением и длительным существованием (Рис. 4). Adocidae были более многочисленны, чем Nanhsiungchelyidae и часто являлись доминантами в позднемеловых комплексах черепах (Несов, Красовская, 1984; Несов, 1997; Hutchison, 2000). Включение в состав адоцид Yehguia из Китая позволяет изменить время появления надсемейства Adocoidea и становления обоих семейств с раннего мела на позднюю юру. В раннем мелу (неокоме) появляются уже несколько различных форм адоцид, среди которых впервые определяются представители Shachemydinae из Японии (Shachemydinae indet. 2, см. Главу 5). Первые Nanhsiungchelyidae появляются в раннем мелу, где они ранее были известны только из баррема/апта Японии и впервые определяются нами из аптаЦальба Монголии. В начале позднего мела (сеноман) Nanhsiungchelyidae, ранее известные только из Японии, впервые выявляются в Монголии и представлены двумя родами (Hanbogdemys иKharakhutulia). Помимо этого подтверждается присутствие Nanhsiungchelyidae в раннем сеномане Средней Азии. В интервале поздний турон - ранний кампан оба семейства достигают своего расцвета: Adocidae представлены двумя родами и семью видами, Nanhsiungchelyidae - двумя родами и пятью видами. На территории Средней Азии и Казахстана Nanhsiungchelyidaeне известны,тогда как Adocidaeпредставлены Adocus и Shachemys, которые зачастую встречаются в одних комплексах. На территории Монголии
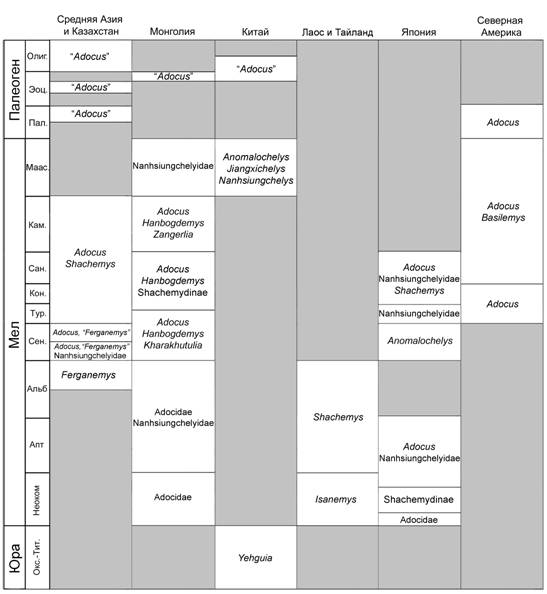
Рис. 4. Геологическое и географическое распространение основных таксонов Adocidae и Nanhsiungchelyidae. Отсутствие находок показано серым цветом. Распространение североамериканских таксонов дано по Hutchison (2000).
этот интервал охарактеризован комплексами верхней части баинширэинской (верхний турон - сантон) и джадохтской (кампан) свит. Из верхов баинширэинской свиты определена единственная пластинка панциря Shachemydinae indet., которая является единственной находкой представителей этого подсемейства в Монголии. По-видимому, в туроне происходит миграция Adocoidea в Северную Америку. В джадохтской свите преобладают остатки Nanhsiungchelyidae, а Adocidae известны слабо и представлены фрагментарными находками Adocussp. На конец позднего мела приходится спад численности Adocoidea: в позднем кампане и маастрихте адоциды уже отсутствуют, а наньсюнгхелииды еще представлены несколькими формами (Anomalochelys, JiangxichelysиNanhsiungchelys). В палеоген переходят только Adocidae, тогда как Nanhsiungchelyidae исчезают в конце мела. Палеогеновые Adocidae (УAdocusФ) малочисленны по сравнению с меловыми и известны по единичным находкам. Первые азиатские УAdocusФ появляются во второй половине палеоцена, что, скорее всего, объясняется их миграцией из Северной Америки (подтверждается особенностями морфологии, см. Главу 3). Наиболее поздние их находки впервые указываются из Казахстана, что продлевает время их существования в Азии с эоцена до конца олигоцена.
6.2. Образ жизни и условия обитания. В результате исследования показано, что обитание Adocidae в основном было приурочено к условиям гумидного субтропического климата на окраинных областях Азии с обширными приморскими низменностями, а Nanhsiungchelyidae - к условиям аридного климата на внутриконтинентальных районах. При этом представители рода Shachemys обитали только в приморских районах, в то время как представители рода Adocus имели широтное распространение, встречаясь как по периферии, так и в центре Азиатского материка.
Вопрос об образе жизни адоцид и наньсюнгхелиид слабо освящен в литературе (Hutchison, Archibald, 1980; Holroyd, Hutchison, 2002). Adocidae рассматриваются как водные черепахи, способные к активному плаванию. Среди них выделяются представители рода Shachemysсхарактерной для них сильной редукцией серии невральных пластинок, что среди черепах известно еще только у некоторых Chelidae (Chelodinaи Emydura). Исчезновение невральных пластинок связано с сокращением пространства для прохождения длиннейшей мышцы спины (m. longissimus dorsi). Слабое развитие этой мышцыограничивает способность к быстрому и резкому выпрямлению шеи, и, таким образом, не позволяет совершать резкие и быстрые выпады за добычей. В связи с этим питание этих черепах сводится к относительно пассивному преследованию и заглатыванию добычи, что можно предполагать и для Shachemys.Вероятно, слабое развитие шейных мышц связано с наличием шарнира в передней части пластрона этих черепах, обеспечивая дополнительную защиту головы при слабо подвижной шее.
Среди Adocidae выявляются различия в пропорциях черепа у Shachemysи Ferganemysс одной стороны и у Adocus - с другой. Так, для Shachemysи Ferganemys характерно наличие невысокого, относительно удлиненного и обтекаемого черепа, тогда как у Adocusон заметно расширен сзади, более высокий и короткий. Различается и положение глазниц - у Shachemysи Ferganemys они смещены наверх, а у Adocus расположены по бокам черепа. Такие различия, по-видимому, связаны с особенностями питания: наличие относительно высокого черепа предполагает большую силу укуса и питание более подвижными объектами для Adocus, чем для Shachemysи Ferganemys.
Для Nanhsiungchelyidae обычно указывается наземный образ жизни, экологически сходный с таковым современных Testudinidae (Hutchison, Archibald, 1986; Brinkman, 2003; Hutchison, Holroyd, 2003). Однако существует и иное мнение, определяющее для них водный образ жизни (Суханов, Нармандах, 1975, 1977; Langston, 1956; Sukhanov, 2000). Полученные нами данные по морфологии Nanhsiungchelyidae (угол между лопаточным и акромиальным отростком более 90?; головка плечевой кости округлая; кости предплечья короткие; подвздошная кость образует более половины поверхности вертлужной впадины; латеральные отростки лобковых костей направлены вбок; кости конечностей укороченные и массивные; фаланговая формула сокращенная) позволяют в целом реконструировать наземный образ жизни для этих черепах. Большинство находок наньсюнгхелиид приурочено к эоловым отложениям джадохтской и кампанской свит, что предполагает их обитание в жарких и засушливых пустынных ландшафтах.
Среди Nanhsiungchelyidae выделяется Nanhsiungchelys ,обладающий относительно крупным черепом с увеличенной носовой частью. Сходное строение известно у Aldabrachelys Loveridge et Williams, 1954, крупные носовые полости которых приспособлены для поглощения воды через ноздри (Arnold, 1979), что является адаптацией к обитанию в засушливых условиях. Возможно, подобное приспособление развилось и у Nanhsiungchelys в условиях аридного климата конца мела.
ВЫВОДЫ
- Для черепах семейств Adocidae и Nanhsiungchelyidae выявлен ряд примитивных признаков морфологии (невральная формула 6>4<6<6<6<6<6, захождение маргинальных щитков на реберные пластинки, наличие высоких задних краевых пластинок, широких долей пластрона, крупных анальных щитков, извилистой срединной борозды пластрона), характерных для базальных эукриптодир уровня Xinjiangchelyidae.
- Показано существование клады Adocusia (неранговый таксон; Adocidae + Nanhsiungchelyidae), которая в ранговой систематике соответствует надсемейству Adocoidea. Эта клада занимает сестринское положение по отношению к кладе Trionychoidea.
- Современная система черепах надсемейства Adocoidea включает 2 семейства (Adocidae и Nanhsiungchelyidae), 2 подсемейства (Adocinae и Shachemydinae), 11 родов, 26 видов, из которых 1 род (Kharakhutulia) и 3 вида (Adocus bostobensis, A. dzhurtasensisи Kharakhutuliakalandadzei) новые для науки. Для 35 форм изменено систематическое положение, описаны 20 новых находок Adocoidea.
- Показано, что Adocidae распространены в Азии в интервале поздняя юра - олигоцен, Shachemydinae выявлены в Монголии в позднем мелу, Nanhsiungchelyidae выявлены в Монголии в раннем мелу, Hanbogdemys распространен в Азии в интервале сеноман - кампан.
- Установлено, что Adocidae были пресноводными черепахами, среди которых выделяются формы с различными пищевыми стратегиями; Nanhsiungchelyidae, вероятно, являлись сухопутными черепахами.
СПИСОК РАБОТ ПО ТЕМЕ ДИССЕРТАЦИИ
Список работ, опубликованных в изданиях рекомендованных ВАК:
1. Danilov I.G., Syromyatnikova E.V. 2008. New materials on turtles of the family Nanhsiungchelyidae from the Cretaceous of Uzbekistan and Mongolia, with a review of the nanhsiungchelyid record in Asia. Proceedings of the Zoological Institute of the Russian Academy of Sciences (Труды Зоологического института РАН), 312(1/2): 3Ц25.
2. Sukhanov V.B., Danilov I.G., Syromyatnikova E.V. 2008. The description and phylogenetic position of a new nanhsiungchelyid turtle from the Late Cretaceous of Mongolia. Acta Palaeontologica Polonica, 53(4): 601Ц614.
3. Syromyatnikova E.V., Danilov I.G. 2009. New material and a revision of turtles of the genus Adocus (Adocidae) from the Late Cretaceous of Middle Asia and Kazakhstan. Proceedings of the Zoological Institute of the Russian Academy of Sciences (Труды Зоологического института РАН), 313(1): 74Ц94.
4. Syromyatnikova E.V. 2011. Turtles of the genus Ferganemys Nessov et Khosatzky, 1977 (Adocoidea: Adocidae): morphology of the shell and phylogenetic position. Proceedings of the Zoological Institute of the Russian Academy of Sciences (Труды Зоологического института РАН), 315(1): 38Ц52.
5. Danilov I.G., Sukhanov V.B., Syromyatnikova E.V. 2011. New materials on turtles of the family Adocidae with a review of the adocid record in Asia. Proceedings of the Zoological Institute of the Russian Academy of Sciences (Труды Зоологического института РАН), 315(2): 101Ц132.
Список работ, опубликованных в других изданиях:
1. Данилов И.Г., Сыромятникова Е.В. 2007. Морфология и систематика черепах рода Shachemysиз Средней Азии. Современная палеонтология: классические и новейшие методы. Третья Всероссийская Научная школа молодых ученых-палеонтологов (3Ц5 октября 2006, Москва, Россия): 21Ц23.
2. Данилов И.Г., Сыромятникова Е.В., Суханов В.Б. 2007. Черепахи рода Shachemys из верхнего мела Азии. Современная палеонтология: классические и новейшие методы - 2007. ПИН РАН, Москва: 63Ц88.
3. Сыромятникова Е.В., Данилов И.Г. 2007. Морфология черепах рода Adocusиз верхнего мела Средней Азии и Казахстана. Современная палеонтология: классические и новейшие методы. Четвертая Всероссийская Научная школа молодых ученых-палеонтологов (15Ц17 октября 2007, Москва, Россия): 48Ц49.
4. Сыромятникова Е.В., Данилов И.Г. 2008. Распространение ископаемых черепах семейств Adocidae и Nanhsiungchelyidae в Азии. Очередная научная сессия по итогам работ 2007 года (8Ц10 апреля 2008, С-Петербург, Россия). Зоологический институт РАН, С-Петербург: 47Ц48.
5. Сыромятникова Е.В., Данилов И.Г. 2008. Филогения и биогеография черепах семейств Adocidae и Nanhsiungchelyidae. Современная палеонтология: классические и новейшие методы. Пятая Всероссийская Научная школа молодых ученых-палеонтологов (6Ц8 октября 2008, Москва, Россия): 50.
6. Danilov I.G., Syromyatnikova E.V. 2009. Phylogeny of the extinct turtle clade Adocusia. Gaffney Turtle Symposium (October 17Ц18, 2009, Royal Tyrrell Museum, Drumheller, Canada). Abstract Volume: 44Ц51.
7. Сыромятникова Е.В., Данилов И.Г., Суханов В.Б. 2009. Морфология и систематическое положение Adocoides аmtgai - черепахи семейства Adocidae из позднего мела Монголии. Палеонтология центральной Азии. Международная конференция к 40-летию совместной Российско-Монгольской палеонтологической экспедиции (18Ц19 ноября 2009, Москва, Россия). ПИН РАН, Москва: 77Ц79.
8. Данилов И.Г., Сыромятникова Е.В. 2009. Филогенетический анализ черепах надсемейства Adocoidea. Современная палеонтология: классические и новейшие методы - 2009. ПИН РАН, Москва: 67Ц82.
9. Сыромятникова Е.В. 2009. Морфология и филогения ископаемых черепах рода Ferganemys. Современная палеонтология: классические и новейшие методы. Шестая Всероссийская Научная школа молодых ученых-палеонтологов (5Ц7 октября 2009, Москва, Россия): 39Ц40.
10. Syromyatnikova E.V., Danilov I.G., Sukhanov V.B. 2011. The morphology and phylogenetic position of Adocus/Adocoides amtgai, an adocid turtle from the Late Cretaceous of Mongolia. SVP 71st Annual Meeting (October 5Ц7, 2009, Las Vegas, USA): 202.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]