
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Воздействие слабых импульсных магнитных полей на перекисное окисление липидов тилакоидных мембран и функционирование фотосинтетической электрон-транспортной цепи в растениях гороха
Автореферат кандидатской диссертации
На правах рукописи
Кальясова Екатерина Андреевна
ВОЗДЕЙСТВИЕ СЛАБЫХ ИМПУЛЬСНЫХ МАГНИТНЫХ ПОЛЕЙ
НА ПЕРЕКИСНОЕ ОКИСЛЕНИЕ ЛИПИДОВ ТИЛАКОИДНЫХ
МЕМБРАН И ФУНКЦИОНИРОВАНИЕ ФОТОСИНТЕТИЧЕСКОЙ
ЭЛЕКТРОН-ТРАНСПОРТНОЙ ЦЕПИ В РАСТЕНИЯХ ГОРОХА
03.01.05 - физиология и биохимия растений
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Пущино - 2012
Работа выполнена на кафедре биохимии и физиологии растений Федерального государственного бюджетного образовательного учреждения высшего профессионального образования Нижегородского государственного университета им. Н.И. Лобачевского
Научный руководитель: доктор биологических наук, профессор
Веселов Александр Павлович
Официальные оппоненты: аа Иванов Борис Николаевич
доктор биологических наук,
Федеральное государственное бюджетное
учреждение науки Институт фундаментальных
проблем биологии РАН, заведующий лабораторией
фотосинтетического электронного транспорта
Паничкин Леонид Александрович
доктор биологических наук, профессор
Российский государственный аграрный аа а университет - МСХА имени К. А. Тимирязева
кафедра физиологии растений, профессор
Ведущая организация: Биолого-почвенный факультет
Санкт-Петербургского государственного университета
Защита состоится ла 5а аа июля 2012 г. в аа11-00а ачасов на заседании диссертационного совета Д 002.066.01 при федеральном государственном бюджетном учреждении науки Институте фундаментальных проблем биологии РАН по адресу: Московская область, г. Пущино, ул. Институтская, д.2, ИФПБ РАН
С диссертацией можно ознакомиться в библиотеке Федерального государственного бюджетного учреждения науки Института фундаментальных проблем биологии РАН
Автореферат разослан л___аа июняаа 2012 г.
 Ученый секретарь
Ученый секретарь
диссертационного совета, аа
кандидат биологических наук аа аа Г.Н. Назарова
![]()
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы.
Несмотря на богатую историю магнитобиологических исследований, насчитывающую более 100 лет, до сих пор существуют трудности с выявлением мишеней и механизмов восприятия магнитных полей (МП) живыми объектами. На роль первичных рецепторов предлагаются жирные кислоты липидов мембран (Новицкая с соавт., 2010; 2008), белковые молекулы (Binhi, Savin, 2002), ионы, находящиеся в активных центрах ферментов (Леднев, 1996), молекулы воды (Дроздов с соавт., 2010); предполагается действие переменных и постоянных магнитных полей на процессы перекисного окисления липидов (ПОЛ) (Пирузян, Аристархов, 2005; Sahebjamei et al., 2007), изменение вязкости мембран (Калипин с соавт., 2005).
Всемирной организацией здравоохранения одним из ключевых направлений будущих исследований для переменных магнитных полей было названо изучение эффектов, опосредованных через взаимодействие с клеточными мембранами (Гигиенические критерииЕ, 1992). Известно, что в растительной клетке в основе ответа на разнообразные внешние воздействия лежит нарушение функционирования фотосинтетической электрон-транспортной цепи (ФЭТЦ), в результате чего происходит изменение генерации всех основных активных форм кислорода (Мерзляк, 1989), которые, в свою очередь способны регулировать практически все важнейшие процессы жизнедеятельности клетки (Тарчевский, 2002), в том числе через модуляцию процессов перекисного окисления липидов мембран.
В свете вышесказанного представляется перспективным изучение эффектов импульсных магнитных полей (ИМП) в системе перекисного окисления липидов тилакоидных мембран, выявление зависимости наблюдаемых изменений от времени воздействия и характеристик поля, выяснение специфичности восприятия МП физиологически отличающимися объектами, а также изучение проявления вызванных магнитным полем изменений ПОЛ в функционировании фотосинтетической электрон-транспортной цепи. Эти знания важны для раскрытия как общебиологических механизмов рецепции магнитных полей, так и выявления особенностей ответа растительных организмов.
Цель и задачи исследования.
Целью данной работы было изучение ответа системы перекисного окисления липидов мембран тилакоидов гороха на воздействие импульсных магнитных полей с различными характеристиками, выявление особенностей функционирования фотосинтетической электрон-транспортной цепи, возникающих в связи с наблюдаемыми изменениями, определение особенностей биохимического ответа у физиологически отличающихся объектов.
Для достижения цели были поставлены следующие задачи:
1.аа Определить содержание продуктов перекисного окисления липидов в тилакоидах, выделенных из растений гороха, обработанных импульсными магнитными полями с различными характеристиками. Выяснить зависимость наблюдаемых изменений от длительности экспозиции растений в поле.
2.аа Выявить особенности функционирования фотосинтетической электрон-транспортной цепи растений, обработанных магнитным полем; проследить их взаимосвязь с изменениями, происходящими в процессе перекисного окисления липидов мембран тилакоидов.
3.аа Оценить стабильность эффектов воздействия импульсного магнитного поля на перекисное окисление липидов тилакоидов гороха.
4.аа Провести сопоставление эффектов и оценить специфичность изменений, происходящих на уровне перекисного окисления липидов тилакоидов при воздействии импульсного магнитного поля на растения гороха различных сортов.
Научная новизна.
Получены новые данные, свидетельствующие о выполнении перекисным окислением липидов мембран тилакоидов роли универсального звена в восприятии растениями низкоинтенсивных импульсных магнитных полей.
Впервые продемонстрировано развитие ответа, зависящего от времени воздействия поля, со стороны фотосинтетической электрон-транспортной цепи.
Исследовано последействие импульсного магнитного поля с частотой 15 Гц и амплитудой 1,5 мТл и показан стабильный характер изменения содержания диеновых конъюгатов и оснований Шиффа - продуктов перекисного окисления липидов мембран хлоропластов.
Проведено сопоставление изменений перекисного окисления липидов, происходящих под действием ИМП, в растениях гороха двух сортов. Показано, что изменения содержания продуктов ПОЛносили универсальный характер.
Теоретическая и практическая значимость.
Показанное в данной работе воздействие ИМП с частотой 15 Гц и амплитудой 1,5 мТл на перекисное окисление липидов тилакоидных мембран, а так же связь наблюдаемых изменений с особенностями функционирования фотосинтетической электрон-транспортной цепи могут иметь большое значение для раскрытия механизмов восприятия растениями низкоинтенсивных магнитных полей. Универсальность ответа со стороны перекисного окисления липидов представляет интерес для понимания общебиологических механизмов магниторецепции.
Полученные данные необходимы для нормирования допустимого воздействия магнитного поля на живые объекты и могут использоваться при биоиндикации и экологическом мониторинге антропогенной нагрузки на фитоценозы.
Апробация работы.
Результаты работы были представлены на II и III Международных конференциях Человек и электромагнитные поля (Саров, 2007, 2010); на I, II и III Всероссийских конгрессах студентов и аспирантов Симбиоз-Россия (Казань, 2008; Пермь, 2009; Н.Новгород, 2010); Международных конференциях по фундаментальным наукам среди студентов и аспирантов Ломоносов (Москва, 2007, 2009, 2010); 12-ой, 14-ой, 15-ой Нижегородских сессиях молодых учёных (Н.Новгород, 2007, 2009, 2010); V международном конгрессе Слабые и сверхслабые поля и излучения в биологии и медицине (Санкт-Петербург, 2009); Всероссийском симпозиуме Растение и стресс (Москва, 2010); 14-ой и 15-ой Пущинских международных школах-конференциях молодых ученых Биология - наука 21го века (Пущино, 2010, 2011); III Международном симпозиуме Клеточная сигнализация у растений (Казань, 2011); VII Съезде Общества физиологов растений Физиология растений - фундаментальная основа экологии и инновационных биотехнологий (Н.Новгород, 2011).
Публикации.
По материалам диссертации опубликовано и направлено в печать 20 работ, в том числе 4 - в рецензируемых изданиях.
Структура и объем диссертации.
Диссертация состоит из введения, обзора литературы, описания методов и объекта исследования, 4 разделов, где представлены результаты исследований и их обсуждение, заключения, выводов, списка литературы и 1 приложения.
ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЙ
Объект исследований и постановка опытов. Для проведения экспериментов использовали 14-дневные растения гороха (Pisum sativum L.) сортов Альбумен или Шустрик, выращенные на фильтровальной бумаге, смачиваемой водопроводной водой, в условиях климатической камеры при 22-23?С.
Сорт Альбумен включен в Госреестр в 1998 году. Стебель обычный, средней длины. Общее число междоузлий 15-19, до первого соцветия 11-14. Лист с 2-3 парами цельнокрайних листочков среднего размера. Прилистники хорошо развиты, имеется пятнистость. Среднеспелый. Устойчивость к полеганию, осыпанию - на уровне стандартных сортов обычного типа.
Сорт Шустрик Включен в Госреестр селекционных достижений РФ в 2003 г., защищен патентом. Безлисточковый, неосыпающийся. Число междоузлий до первого соцветия 9-10. Прилистники хорошо развиты, имеется пятнистость. Раннеспелый. Низкорослый - 40-65 см.
Исследования проводились в нескольких вариантах:
1. Целые растения обрабатывали импульсным магнитным полем, с индукцией 3,5 мТл и частотой 100 Гц (ИМП-1), создаваемым с помощью магнитотерапевтической установки УМТИ-3Ф Колибри. В качестве объекта использовали растения гороха сорта Альбумен. Выделение тилакоидов и анализ проводили сразу после экспозиции в поле, длительностью 15, 30, 60 и 120 минут.
2. Целые растения обрабатывали импульсным магнитным полем (ИМП-2) - пачки из 20 импульсов длительностью 227 мкс с амплитудой 1,5 мТл, следующих с частотой 15 Гц, для создания поля использовали генератор фирмы Electro-Biology Inc. (EBI). В качестве объекта использовали растения гороха сорта Альбумен. Растения подвергали обработке полем в течение 15, 30, 60 и 120 минут. Тилакоиды выделяли как непосредственно после обработки полем, так и через 30 минут после окончания воздействия.
3. В качестве объекта использовали растения гороха сорта Шустрик. Целые растения обрабатывали ИМП-2. Сразу после окончания экспозицииа (15, 30, 60 и 120 минут) из растений получали суспензию тилакоидов, которую и использовали для проведения анализа.
Во всех опытных постановках контролем служила суспензия тилакоидов из растений, не подвергнутых перечисленным воздействиям.
Получение суспензии тилакоидов. Исследования проводили на суспензии тилакоидов, изолированных по методу дифференциального центрифугирования (Гавриленко, Жигалова, 2003).
Определение содержания продуктов липопероксидации. Развитие перекисного окисления липидов оценивали по уровню первичных (диеновые конъюгаты и липидные гидроперекиси) и вторичных (малоновый диальдегид и основания Шиффа) продуктов спектрофотометрически (Стальная, Гаришвили, 1977; Романова, Стальная, 1977; Волчегорский с соавт., 1989). Общее количество липидов определяли спектрофотометрически по реакции с фосфорно-ванилиновым реактивом (Камышников, 2000).
Определение фотосинтетической активности. Проводили на целых растениях методом индукции флуоресценции хлорофилла (ИФХ) (Корнеев, 2002; Schreiber, Klughammer, 2008; Klughammer, Schreiber, 2008) с использованием прибора Dual-PAM-100 (Walz, Германия).
Параметры освещения:
ML (measuring light) - измеряющий свет (плотность потока фотонов 24 мкмоль фотонов*м-2*с-1; ?=460 нм);
AL (acting light) - действующий (актиничный) свет, (плотность потока фотонов 103 мкмоль фотонов*м-2*с-1; ?=460 нм);
SP (saturation pulses) - короткие вспышки насыщающего света, (плотность потока фотонов 10000 мкмоль фотонов*м-2*с-1, длительность импульсов 300 мс; ?=635 нм; частота вспышек 5 с).
Статистическая обработка полученных результатов. Обработку полученных результатов производили методами параметрической статистики с определением стандартной ошибки (Гланц, 1998). На рисунках представлены средние арифметические 3-6 независимых опытов, каждый из которых проводили в трехкратной биологической повторности. Значимость различий оценивали по критерию Стьюдента с поправкой Бонферрони.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
1. Воздействие импульсных магнитных полей на перекисное окисление липидов тилакоидов гороха сорта Альбумен.
1.1. Воздействие ИМП-1.
Биологические мембраны неоднократно предлагались на роль неспецифических сенсоров различных физических воздействий (Эйдус, 2001; Курганова с соавт., 1997), в том числе и магнитного поля (Пономарев, Новиков, 2009). Содержание продуктов ПОЛ напрямую влияет на целостность и функционирование мембран.
В наших экспериментах магнитное поле с частотой колебаний 100 Гц, амплитудой 3,5 мТл (ИМП-1) с 30 по 60 мин воздействия снижало содержание продуктов перекисного окисления липидов (рис.1), а именно, уровень диеновых конъюгатов (ДК) и малонового диальдегида (МДА), был минимальным после 60-минутной экспозиции и составил 64 % от контроля (2,16 мкмоль/мг липидов) и 38 % от контроля (37 мкмоль/мг липидов) соответственно. К 120-мин экспозиции эти показатели несколько возрастали, но так и не достигали контрольного уровня.
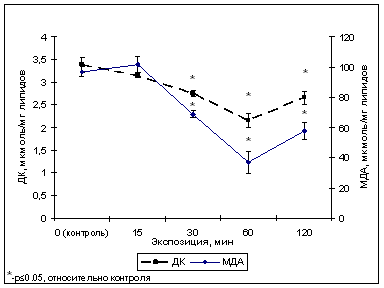
Рис. 1. Содержание продуктов ПОЛ в тилакоидах гороха сорта Альбумен после обработки ИМП-1.
Говоря о возможных путях воздействия магнитных полей на мембраны, приводящих к отмеченным изменениям, можно выделить их влияние на выход конечных продуктов в реакциях перекисного окисления ненасыщенных жирных кислот фосфолипидов мембран с участием свободных радикалов, основанную на существовании механизма влияния МП на скорость рекомбинации радикальных пар (Пирузян, Аристархов, 2005). В экспериментах с использованием растительных объектов отмечено изменение кинетики проницаемости мембран для ионов в результате действия магнитного поля (Новицкая с соавт., 2006), что так же может быть связано с состоянием липидного компонента мембран.
Таким образом, результатом воздействия ИМП-1 стало уменьшение содержания продуктов липопероксидации в мембранах хлоропластов, которое могло происходить вследствие нарушения полем рекомбинации радикальных пар, образующихся в процессе ПОЛ.
1.2. Воздействие ИМП-2.
По своим частотным характеристикам ИМП-2 (15 Гц), как и ИМП-1 (100 Гц) относится к низкочастотным полям, для которых характерно нетепловое воздействие на биосистемы. Частоты исследуемых полей совпадают с диапазоном низкочастотных пульсаций магнитного поля Земли (0,1-100 Гц), наибольшая амплитуда которых приходится на частоты 8-16 Гц (Петин, 2006).
В наших экспериментах 15-минутная обработка ИМП-2 не вызывала изменения содержания продуктов липопероксидации - ДК и оснований Шиффа (ОШ), тогда как 30- и 60-минутные воздействия приводили к ингибированию процессов ПОЛ. Уровень ДК после 30-минутной обработки составлял 1,65 мкмоль/мг липидов (82 % от контрольного), после 60-минутной - 1,5 мкмоль/мг липидов (81 % от контрольного) (рис.2а), содержание ОШ после 30-минутной обработки составило 7,86 ед.опт.пл/мг липидов (81 % от контрольного уровня), после 60-минутной - 7,48 ед.опт.пл/мг липидов (91 % от контроля) (рис.2б). Более длительная 120-минутная обработка не вызывала значимого изменения уровня продуктов ПОЛ.
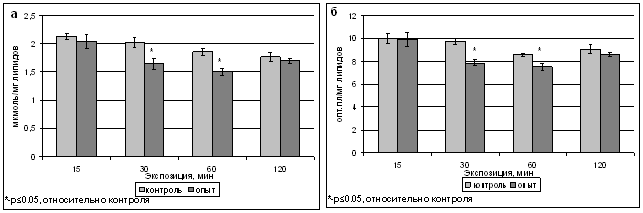
Рис. 2. Содержание продуктов ПОЛ - диеновых конъюгатов (а) и оснований Шиффа (б) в тилакоидах гороха сорта Альбумен после обработки ИМП-2.
Для развития процесса ПОЛ и нарушения барьерных свойств мембран существенную роль играет накопление липидных гидроперекисей (ГП). В наших экспериментах происходило накопление ГП после 60- и 120-минутной обработок, до 121 % и 146 % от контроля (2,24 и 2,02 ед.опт.пл/мг липидов) соответственно, менее длительные обработки не вызывали изменения данного параметра (рис.3).
Таким образом, если содержание продуктов ПОЛ на протяжении всех времен обработки не превышало контрольного уровня или даже оказывалась ниже его, то длительные (60 и 120 минут) обработки растений полем приводили к динамическому накоплению ГП.
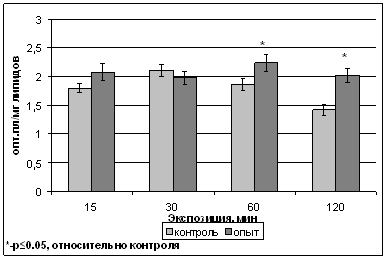
Рис. 3. Содержание липидных гидроперекисей в тилакоидах гороха сорта Альбумен после обработки ИМП-2.
Увеличение содержания гидроперекисей могло быть связано с накоплением АФК. Так, например, при экспонировании ФМА - активированных нейтрофилов в поле, аналогичном ИМП-2 наблюдалось увеличение скорости генерации АФК, регистрируемой по люминол - зависимой хемилюминесценции на 25-30 % (Белова, 2001). В наших экспериментах накопление АФК, в частности, пероксида водорода, могло происходить в результате разбалансировки окислительно-восстановительных процессов, протекающих на тилакоидных мембранах.
Сопоставляя результаты изучения изменения уровня продуктов ПОЛ в результате действия ИМП-1 с данными, полученными при использовании ИМП-2 (менее мощного поля), можно говорить об идентичности характера ответа со стороны окислительных процессов в мембранах тилакоидов гороха. Причем, амплитуда наблюдаемых изменений находилась в прямой зависимости от интенсивности воздействия - более мощное поле вызывало более заметные изменения. Менее интенсивное поле, по-видимому, производило меньшие возмущения в этих системах вне зависимости от времени воздействия. Отличия в проявлении биоэффектов ИМП-1 и ИМП-2 могли быть обусловлены различными механизмами их возникновения. Так для ИМП-2 (частота 15 Гц),а наиболее значимым является резонансное влияние поля на биомолекулы и биологические процессы, а для ИМП-1 (частота 100 Гц) подобный механизм маловероятен (Узденский, 2000). В то же время, с увеличением частоты поля увеличивается его воздействие на рекомбинацию радикальных пар, что может сопровождаться нарушением свободнорадикальных процессов, активацией антиоксидантной системы и увеличением амплитуды изменения наблюдаемых показателей.
2. Особенности функционирования фотосинтетической электрон-транспортной цепи растений гороха сорта Альбумен, обработанных импульсным магнитным полем.
Фотосистема II (ФС II) играет центральную роль в осуществлении и регуляции электронного транспорта в хлоропластах. Поэтому показатели, отражающие эффективность работы ФС II являются важным компонентом мониторинга активности фотосинтетического аппарата.
В наших экспериментах наиболее показательные изменения индукционных кривых наблюдались в течение первых 2 минут после начала освещения. В результате действия МП происходило изменение квантового выхода ФС II (рис. 4, а-г). 15-минутная обработка не вызывала изменений данного параметра, тогда как 30-минутная приводила к некоторому увеличению квантового выхода ФС II, который после 60-минутной обработки возвращался к контрольным значениям и несколько снижался после 120-минутной обработки. Следует отметить, что 30- и 60-минутная обработки полем приводили к более быстрому достижению максимальных значений квантового выхода.
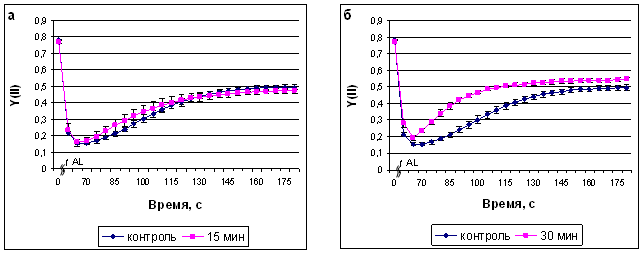
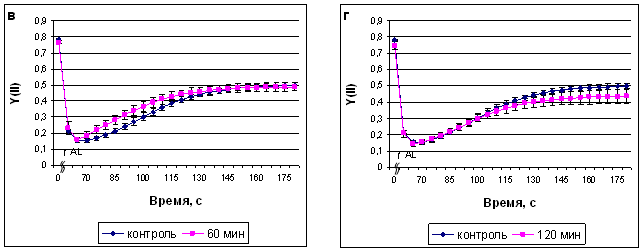
Рис. 4. Квантовый выход ФС II после экспозиции растений сорта Альбумен в ИМП-2, длительностью 15 мин (а), 30 мин (б), 60 мин (в) и 120 мин (г). Начало оси времени совпадает с включением измеряющего света, ^ AL - включение действующего света.
Важной характеристикой состояния ФС II является коэффициент фотохимического тушения, характеризующий количество "открытых" реакционных центров. Коэффициент фотохимичекого тушения показывает долю комплексов ФС II с окисленным акцептором до применения вспышки насыщающего света (Schereiber et al., 1986). В наших экспериментах, колебания данного показателя под действием ИМП-2 совпадали с изменениями квантового выхода ФС II. Увеличение фотохимического тушения флуоресценции хлорофилла происходит при активации процессов, поддерживающих электронный транспорт, в первую очередь, при ускорении фиксации углекислого газа. Поэтому, увеличение коэффициента фотохимического тушения, наблюдавшееся нами после 30-минутной обработки растений МП, может быть связано с активацией реакций, потребляющих АТФ и НАДФН, так как при этом ускоряется окисление компонентов цепи электронного транспорта и отток электронов от QA (Корнеев, 2002).
Другим процессом, влияющим на квантовый выход ФС II является тепловая диссипация энергии поглощенных квантов света, с величиной которой связан коэффициент нефотохимического тушения флуоресценции хлорофилла. Колебания данного параметра отражают регуляторные изменения тепловой диссипации в комплексах ФС II. В наших экспериментах 15-минутная обработка полем не вызывала изменения данного параметра, 30-минутная приводила к более быстрому росту данного параметра, по сравнению с контролем, однако уже через 1 мин после начала наблюдения коэффициент нефотохимического тушения оказывался меньше контрольного уровня, что, по-видимому, и обеспечивало рост квантового выхода и коэффициента фотохимического тушения. 60-минутная обработка полем так же приводила к более быстрому достижению максимального значения данного параметра, наблюдалась тенденция к увеличению его стационарного уровня. Значения коэффициента нефотохимического тушения после 120-минутной обработки ИМП-2 не отличались от контроля.
Нефотохимическое тушение зависит от различных процессов и состоит из нескольких составляющих. Существуют несколько компонентов нефотохимического тушения, различающихся по скорости темновой релаксации (Horton, Hague, 1988; Walters, Horton, 1993; Maxwell, Jonson, 2000). Наблюдавшиеся нами изменения коэффициента нефотохимического тушения, после обработок растений МП в течение 30 и 60 минут, вероятно, были связаны с его быстрым компонентом, который зависит от трансмембранного градиента протонов и модулируется степенью деэпоксидации пигментов виолоксантинового цикла, непосредственным участником которого является аскорбат (Alsher et al., 1997).
Изменения квантового выхода фотосистемы I (ФС I) носили схожий характер с изменениями квантового выхода ФС II (рис.5, а-г).
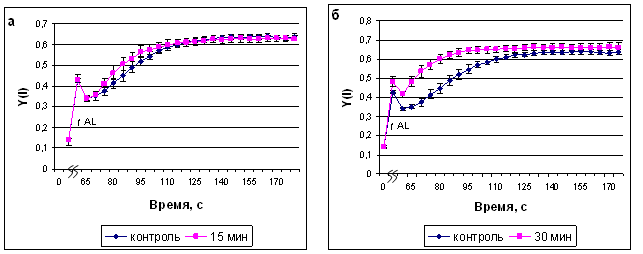
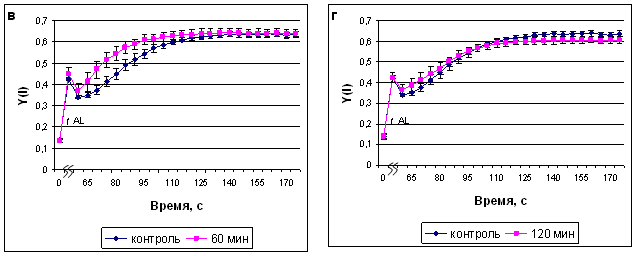
Рис. 5. Квантовый выход ФС I после экспозиции растений сорта Альбумен в ИМП-2, длительностью 15 мин (а), 30 мин (б), 60 мин (в) и 120 мин (г). Начало оси времени совпадает с включением измеряющего света, ^ AL - включение действующего света.
С квантовым выходом ФС I связаны значения показателей нефотохимической диссипации энергии на донорной (Y(ND)) и акцепторной (Y(NA)) сторонах фотосистемы I.
В наших экспериментах эти показатели изменялись соответственно колебаниям квантового выхода ФС I, но наиболее выраженными были изменения на донорной стороне реакционных центров. 15- минутная обработка полем не влияла на диссипацию энергии на донорной стороне ФС I; 30-минутная обработка приводила к более быстрому достижению максимальной окисленности донорной стороны, при этом на второй минуте после насыщающей вспышки значение Y(ND) стабилизировалось на уровне, меньшем контрольного, 60-минутная обработка также вызывала более быструю стабилизацию данного параметра, но в дальнейшем этот показатель не отличался от контрольного. 120-минутная обработка полем существенно не меняла характер ответа, но приводила к повышению уровня нефотохимической диссипации энергии на донорной стороне на протяжении всего времени наблюдения.
Таким образом, 30-минутная обработка растений ИМП-2 вызывала уменьшение содержания ДК и ОШ, что должно было приводить к увеличению гидрофобности тилакоидных мембран и изменению их проницаемости для ионов. Изменение свойств мембран, в свою очередь, приводило к уменьшению нефотохимического тушения, и, как следствие, к увеличению квантового выхода ФС II. Увеличение квантового выхода ФС I было связано с уменьшением тепловой диссипации энергии на ее донорной стороне, также вызванном изменением состояния мембран. Нарушение ионной проницаемости тилакоидных мембран могло отразиться на синтезе АТФ и оказывать влияние на темновую фазу фотосинтеза.
60-минутная экспозиция, помимо снижения содержания ДК и ОШ сопровождалась накоплением липидных гидроперекисей, что, по-видимому, в какой-то степени нормализовывало состояние мембран и поддерживало функционирование ФЭТ - на контрольном уровне. 120-минутная обработка не только вызвала еще большее накопление гидроперекисей липидов, но и сопровождалась увеличением содержания других продуктов ПОЛ - ДК и ОШ до контрольного уровня, что усилило окислительную модификацию мембран, увеличило нефотохимическое тушение в ФС II и диссипацию энергии на донорной стороне ФС I и отрицательно сказалось на работе ФЭТ - приводя к некоторому снижению квантовых выходов обеих фотоситем.
3. Оценка стабильности изменений содержания продуктов перекисного окисления липидов, вызванных воздействием ИМП-2, в тилакоидах гороха сорта Альбумен.
Для оценки стабильности наблюдаемых изменений содержания продуктов перекисного окисления липидов проводили измерение исследуемых параметров через 30 минут после окончания соответствующей обработки полем.
Через 30 мин после 15-минутной обработки наблюдали снижение уровня ДК до 1,87 мкмоль/мг липидов (88 % от контроля), что говорило об активации защитных систем даже после столь короткой экспозиции в поле. Содержание ДК через 30 минут после остальных экспозиций значимо не отличалось от наблюдаемого сразу после окончания обработки (рис.6а).
Содержание ОШ так же практически не изменялось по сравнению с уровнем, наблюдавшимся сразу после окончания обработки полем (рис.6б), лишь только через 30 минут после окончания 30-минутной экспозиции в поле наблюдали некоторый подъем уровня ОШ относительно контроля.
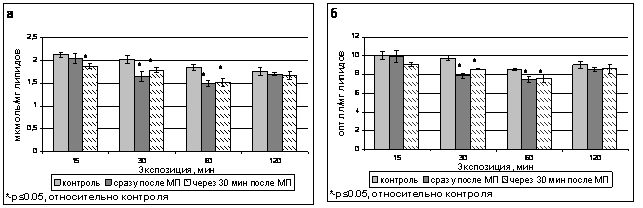
Рис. 6. Содержание продуктов ПОЛ - ДК (а) и ОШ (б) в тилакоидах гороха сорта Альбумен сразу после обработки ИМП-2 и через 30 мин после ее окончания.
Таким образом, изменение содержания ДК и ОШ под воздействием магнитного поля носило стойкий характер.
В отличие от ДК и ОШ, для липидных гидроперекисей был продемонстрирован скоротечный характер наблюдаемых изменений - через 30 минут после окончания воздействия уровень гидроперекисей возвращался к контрольному (рис.7).
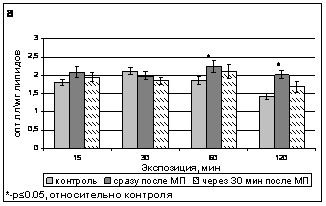
Рис. 7. Содержание ГП в тилакоидах гороха сорта Альбумен сразу после обработки ИМП-2 и через 30 мин после ее окончания.
Наблюдавшиеся нами изменения содержания продуктов ПОЛ могли быть как результатом прямого воздействия поля, так и свидетельствовать об изменении со стороны антиоксидантной системы.
Образование липидных гидропероксидов в процессе ПОЛ происходит при активном участии токоферола, восстановление которого, в свою очередь сопровождается расходованием аскорбата и переходом его в монодегидроаскорбат (Munne-Bosch, 2005). В то же время, образующиеся в качестве промежуточных продуктов ПОЛ гидропероксиды являются высокогидрофильными веществами, они эффективно разрыхляют бислой, увеличивая доступность гидрофобных доменов для воды (Каган, Орлов, Пименко, 1986), что естественно нарушает как активные, так и пассивные процессы перемещения веществ через мембраны, а, следовательно, может оказывать влияние на функционирование антиоксидантной системы. Быстрая нормализация уровня липидных гидроперекисей в течение 30 минут после окончания обработки растений магнитным полем может объясняться непосредственным влиянием ИМП на содержание ГП, с прекращением действия поля исчезал и фактор, вызывающий их накопление. Изменение содержания продуктов ПОЛ влияет на состояние мембран, что, в свою очередь модифицирует фотосинтетическую активность хлоропластов.
4. Воздействие ИМП-2 на перекисное окисление липидов в тилакоидах гороха сорта Шустрик.
Наблюдение магнитобиологических эффектов и их стабильность обусловлены одновременным попаданием в электромагнитные (частотные и амплитудные), временные и физиологические окна (Hajnorouz et al., 2011; Бинги, Савин, 2003). Следовательно, получение репрезентативных результатов при использовании физиологически гетерогенных объектов становится особенно актуальным. С этой целью нами была проведена серия экспериментов с использованием ИМП-2, где в качестве объекта выступал горох сорта Шустрик - безлисточковый, с хорошо развитыми прилистниками. Показано, что безлисточковые формы уступают листочковым по комплексу показателей водного обмена (Новикова, 2009), что от морфологических и биологических особенностей сорта зависит и антиоксидантная активность (Друшляк, 2008).
В наших экспериментах с использованием растений гороха сорта Шустрик, 15-60 минутные обработки ИМП-2 не изменяли уровень диеновых конъюгатов, тогда как после 120- минутной обработки содержание ДК падало на 22 % относительно контрольного уровня и составляло 3,08 мкмоль/мг липидов (рис.8а). В то же время, при использовании в качестве объекта растений сорта Альбумен наблюдалось более раннее снижение содержания ДК - после 30- и 60-минутных обработок полем (рис.8б).
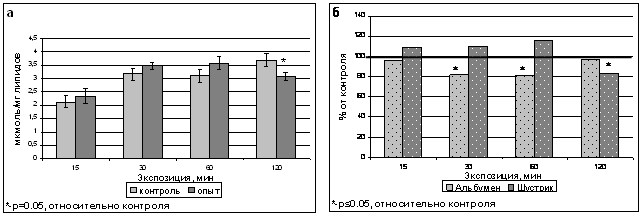 Рис. 8. Содержание диеновых конъюгатов в тилакоидах гороха сорта Шустрик после обработки ИМП-2 (а) и сравнение содержания ДК в тилакоидах гороха сортов Альбумен и Шустрик после обработки магнитным полем (б).
Рис. 8. Содержание диеновых конъюгатов в тилакоидах гороха сорта Шустрик после обработки ИМП-2 (а) и сравнение содержания ДК в тилакоидах гороха сортов Альбумен и Шустрик после обработки магнитным полем (б).
Содержание ОШ, также как и ДК в растениях гороха сорта Шустрик не изменялось после 15-60 минутных обработок полем, снижалось после 120-минутной экспозиции на 17% относительно контроля (рис.9а). После 30- и 60-минутных обработок магнитным полем гороха сорта Альбумен наблюдалось более раннее снижение содержания ОШ, а 120-минутное воздействие не влияло на данный параметр (рис.9б).
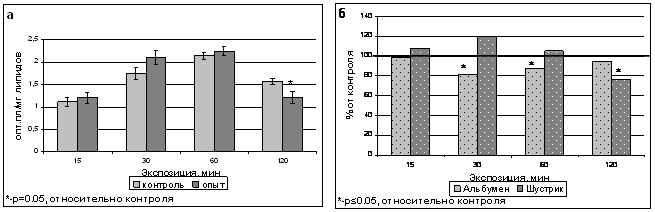
Рис. 9. Содержание оснований Шиффа в тилакоидах гороха сорта Шустрик после обработки ИМП-2 (а) и сравнение содержания ОШ в тилакоидах гороха сортов Альбумен и Шустрик после обработки магнитным полем (б).
Снижение уровня продуктов ПОЛ может происходить в результате непосредственного воздействия МП на процесс липопероксидации. Однонаправленное изменение содержания продуктов перекисного окисления липидов в результате воздействия ИМП на растения гороха обоих сортов может говорить о схожем исходном физико-химическом состоянии фосфолипидов мембран тилакоидов, так как именно от их исходного состояния зависит направление изменений (ускорение или торможение) процессов ПОЛ, вызванных магнитным полем (Пирузян, Аристархов, 2005). Более позднее снижение содержания продуктов ПОЛ в тилакоидах безлисточкового гороха свидетельствует о большей устойчивости растений сорта Шустрик к воздействию слабого импульсного магнитного поля и может быть связано с особенностями липидного состава фотосинтетических мембран.
ЗАКЛЮЧЕНИЕ
Полученные результаты свидетельствуют о том, что возникновение ответа на действие импульсных магнитных полей со стороны перекисного окисления липидов тилакоидов гороха носило неспецифический характер и не зависело ни от интенсивности поля, ни от сортовой принадлежности объекта исследования. Длительность обработки растений магнитным полем, которая вызывала изменения со стороны продуктов ПОЛ, зависела от морфофизиологических особенностей объекта, а сила (амплитуда) ответа была связана с интенсивностью действующего поля.
ИМП низкой интенсивности вызывало дозозависимые изменения со стороны обеих фотосистем: 15-минутная обработка полем не вызывала значимых изменений; повышение квантовых выходов после 30-минутной обработки полем сменялось переходным состоянием мембран, а затем наблюдалось некоторое ингибирование световой фазы фотосинтеза после 120-минутной экспозиции растений в поле, совпадавшее с активацией окислительных процессов на мембранах тилакоидов. Схема возможного развития обсуждаемых процессов представлена на рис.11.
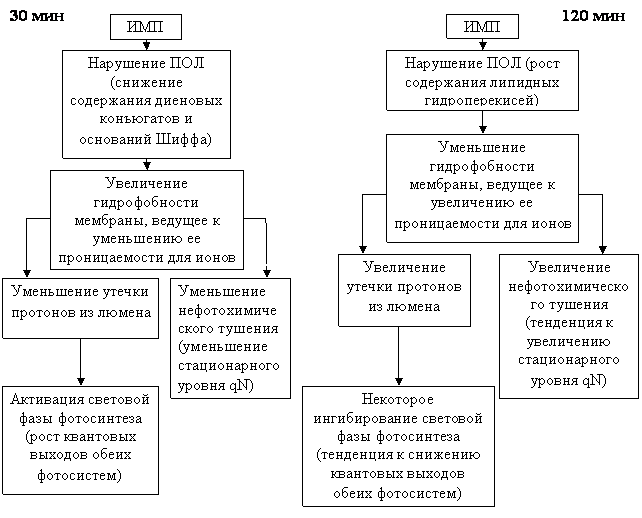
Рис.11. Схема возможной взаимосвязи изменений ПОЛ мембран тилакоидов и особенностей функционирования фотосинтетической электрон-транспортной цепи растений гороха, обработанных магнитным полем.
Полученные данные указывают на влияние изменений, вызванных импульсным магнитным полем в перекисном окислении липидов тилакоидных мембран на функционирование фотосинтетической электрон-транспортной цепи.
ВЫВОДЫ
- Действие ИМП-1 (100 Гц, 3,5 мТл) и ИМП-2 (15 Гц, 1,5 мТл) на процессы ПОЛ носило схожий дозозависимый характер - 15-минутная обработка не вызывала изменений со стороны продуктов ПОЛ; 30- и 60-минутная обработки приводили к снижению, а 120-минутная - к возрастанию их уровня до контрольных значений. Амплитуда наблюдаемых изменений находилась в прямой зависимости от интенсивности воздействия.
- Под действием ИМП-2 после 60- и 120-минутной обработки наблюдалось накопление липидных гидроперекисей в тилакоидных мембранах.
- Изменение ПОЛ тилакоидных мембран приводило к модификации фотосинтетического электронного транспорта. Со стороны фотосинтетических процессов наблюдалось увеличение квантового выхода ФС I после 30-минутной обработки полем, вызываемое уменьшением нефотохимического тушения на донорной стороне реакционных центров, и увеличение квантового выхода ФС II, связанное с уменьшение нефотохимического тушения, что говорит об активации всей электрон-транспортной цепи. 120-минутная обработка приводила к некоторому снижению квантовых выходов обеих фотосистем, что в совокупности с накоплением липидных гидроперекисей свидетельствует о начале повреждения мембран и ингибирования фотосинтетических процессов.
- Обработка ИМП-2 имела эффект последействия. Через 30 минут после окончания экспозиций сохранялись изменения содержания продуктов ПОЛ - диеновых конъюгатов и оснований Шиффа, при этом уровень липидных гидроперекисей снижался до контрольного.
- Процесс перекисного окисления липидов безлисточкового сорта гороха проявил меньшую восприимчивость к ИМП по сравнению с аналогичным параметром сорта с обычной морфологией листа, что приводило к более медленному снижению уровня продуктов ПОЛ, наблюдаемому лишь после 120-минутной экспозиции растений в поле.
СПИСОК ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРТАЦИИ
Работы, опубликованные в ведущих рецензируемых научных журналах и изданиях, рекомендуемых ВАК:
- Васильева Е.А., Синицына Ю.В., Половинкина Е.О., Веселов А.П., Цыганова М.И. Изменение некоторых параметров перекисного гомеостаза хлоропластов гороха в ответ на действие физических факторов низкой интенсивности // Вестник Нижегородского университета им. Н.И. Лобачевского - Н.Новгород, №2. Ч.2. Изд-во ННГУ им. Н.И. Лобачевского. 2010. С. 498-503.
- Васильева Е.А., Синицына Ю.В., Веселов А.П, Курганова Л.Н., Балалаева И.В., Цыганова М.И. Прооксидантно-антиоксидантный статус хлоропластов гороха при действии стрессирующих абиотических факторов среды/ 1. Продукция активных форм кислорода и липопероксидация // Вестник Нижегородского университета им. Н.И. Лобачевского - Н.Новгород, №2. Ч.2. Изд-во ННГУ им. Н.И. Лобачевского. 2010. С. 544-549.
- Васильева Е.А., Яшина Е.С., Половинкина Е.О., Синицына Ю.В., Веселов А.П. Влияние импульсных магнитных полей различных характеристик на некоторые компоненты перекисного гомеостаза хлоропластов гороха // Вестник Нижегородского университета им. Н.И. Лобачевского - Н.Новгород, №2. Ч.2. Изд-во ННГУ им. Н.И. Лобачевского. 2011. С. 174-177.
- Половинкина Е.О., Кальясова Е.А., Синицына Ю.В., Веселов А.П. Изменение уровня перекисного окисления липидов и активности компонентов антиоксидантного комплекса в хлоропластах гороха при воздействии слабых импульсных магнитных полей // Физиология растений. 2011. Т.58. №6. С.930-934.
Статьи, доклады, тезисы докладов региональных и международных конференций:
- Половинкина Е.О., Васильева Е.А. Модуляция перекисного гомеостаза мембран хлоропластов гороха переменным магнитным полем // Материалы Международной конференции по фундаментальным наукам среди студентов и аспирантов Ломоносов-2007. - Москва, МГУ. 2007. С. 271.
- Васильева Е.А., Половинкина Е.О., Быкова О.В. Влияние физических факторов - ионизирующего излучения и низкочастотного переменного магнитного поля в малых дозах на прооксидантно-антиоксидантную систему хлоропластов гороха // Материалы 60-й юбилейной научной студенческой конференции биологического факультета ННГУ Биосистемы: организация, поведение и управление. - Нижний Новгород, ННГУ. 2007. С.13-14.
- Половинкина Е.О., Васильева Е.А., Быкова О.В. Изменение перекисного гомеостаза мембран хлоропластов гороха в связи с действием низкочастотного переменного магнитного поля и ионизирующей радиации в малых дозах // Материалы XII Нижегородской сессии молодых ученых Естественные дисциплины. - Нижний Новгород. 2007. С. 20-21.
- Половинкина Е.О., Васильева Е.А., Курганова Л.Н. Модуляция перекисного гомеостаза мембран хлоропластов гороха переменным магнитным полем // Человек и электромагнитные поля. Сборник материалов докладов II Международной конференции: - Саров: РФЯЦ-ВНИИЭФ. 2008. С.73-77.
- Потехина Е.С., Васильева Е.А., Зинина С.В., Синицына Ю.В., Половинкина Е.О. Изменение активности реакции Хилла и некоторых показателей прооксидантно-антиоксидантного равновесия в хлоропластах гороха (PisumsativumL.) при облучении и гипертермии // Биология: традиции и инновации в XXI веке. Материалы I Всероссийского конгресса студентов и аспирантов-биологов Симбиоз-Россия-2008:-Казань:Изд-во Казанск. гос. ун-та. 2008. С.91-93.
- Васильева Е.А. Динамика генерации продуктов перекисного окисления мембран хлоропластов гороха PisumsativumL.в ответ на действие физических факторов различной природы // Ломоносов-2009: Международная конференция студентов, аспирантов и молодых ученых: Тезисы докладов:-М.:Макс пресс. 2009. С.226.
- Знобищева А.В., Сребницкая Л.К., Васильева Е.А., Белова Н.А Влияние крайне слабых магнитных полей на продукты перекисного окисления липидов в хлоропластах гороха // V международный конгресс Слабые и сверхслабые поля и излучения в биологии и медицине, тезисы докладов. 2009. С.81.
- Васильева Е.А., Синицына Ю.В., Половинкина Е.О. Изменение активности антиоксидантных ферментов и содержания продуктов перекисного окисления липидов в хлоропластах гороха под действием низкоинтенсивного магнитного поля // Симбиоз Россия 2009: материалы II Всероссийского с международным участием конгресса студентов и аспирантов-биологов:-Перм.гос.ун-т.-Пермь. 2009. С.95-97.
- Васильева Е.А., Яшина Е.С. Влияние низкоинтенсивного импульсного магнитного поля на перекисный гомеостаз зеленых клеток гороха // Ломоносов-2010: Международная конференция студентов, аспирантов и молодых ученых, Москва, Тезисы докладов, М.: МАКС Пресс. 2010. С. 248-249.
- Васильева Е.А., Яшина Е.С., Богатырева Е.А. Влияние импульсного магнитного поля на перекисный гомеостаз хлоропластов гороха // Биология-наука 21 века: 14-я Пущинская международная школа-конференция молодых ученых, сборник тезисов, Пущино. 2010. Т.2. С.312-313.
- Васильева Е. А., Яшина Е.С. Влияние импульсного магнитного поля на перекисный гомеостаз хлоропластов гороха // Сборник тезисов III Всероссийского c международным участием конгресса аспирантов и студентов - биологов Симбиоз Россия - 2010. - Нижний Новгород. 2010. С. 47
- Васильева Е.А., Яшина Е.С., Половинкина Е.О., Синицына Ю.В. Изменение компонентов перекисного гомеостаза зеленых клеток гороха в ответ на действие импульсного магнитного поля // Человек и электромагнитные поля. Сборник материалов докладов III Международной конференции: - Саров: РФЯЦ-ВНИИЭФ. 2010. С.125-129.
- Васильева Е.А., Веселов А.П., Половинкина Е.О., Синицына Ю.В. Изменение параметров перекисного гомеостаза хлоропластов гороха в связи с действием переменных магнитных полей с различными характеристиками // Тезисы докладов Всероссийского симпозиума Растение и стресс, Москва. 2010. С. 78-79.
- Кальясова Е.А., Яшина Е.С. Воздействие импульсного магнитного поля на прооксидантно-антиоксидантный статус хлоропластов гороха // Биология-наука 21 века: 15-я Пущинская международная школа-конференция молодых ученых, Пущино. 2011. С.420.
- Кальясова Е.А., Половинкина Е.О., Синицына Ю.В., Веселов А.П Воздействие низкочастотных импульсных магнитных полей с различными характеристиками на перекисный гомеостаз хлоропластов гороха // III Международный симпозиум Клеточная сигнализация у растений, Казань, 2011, в печати.
- Кальясова Е.А., Половинкина Е.О., Синицына Ю.В., Веселов А.П. Воздействие слабого магнитного поля на некоторые компоненты перекисного гомеостаза клеток гороха // VII Съезд Общества физиологов растений России "Физиология растений - фундаментальная основа экологии и инновационных биотехнологий" и Международная научная школа "Инновации в биологии для развития биоиндустрии сельскохозяйственной продукции". Материалы докладов (в двух частях). Ч.1. - Нижний Новгород. 2011. С.305-306.
СПИСОК ПРИНЯТЫХ СОКРАЩЕНИЙ
АФК - активные формы кислорода
ДК - диеновые конъюгаты
МДА - малоновый диальдегид
ОШ - основания Шиффа
ГП - гидроперекиси липидов
МП - магнитное поле
ИМП - импульсное магнитное поле
ИМП 1 - импульсное магнитное поле первого типа (частота 100 Гц, амплитуда 3,5 мТл)
ИМП 2 - импульсное магнитное поле второго типа (частота 15 Гц, амплитуда 1,5 мТл)
ПОЛ - перекисное окисление липидов
ФЭТ - - фотосинтетическая электрон-транспортная цепь
ФС I, II - фотосистема I, II
ИФХ - индукция флуоресценции хлорофилла
Y(ND) - нефотохимическая диссипация энергии на донорной стороне ФС I
Y(NA) - нефотохимическая диссипация энергии, на акцепторной стороне ФС I
Подписано в печать 1.06.2012. Формат 60Х84 1/16.
Бумага офсетная. Печать офсетная.
Усл. печ. л. 1,5. Тир. 100. Зак.
Типография Нижегородского госуниверситета.
Лицензия № 18-0099.
603000, Н.Новгород, ул. Б. Покровская, 37.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]