
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
ДИНАМИКА КЛЕТОЧНОГО ДЫХАНИЯ И СОДЕРЖАНИЕ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ В ВОЛОКНАХ КАМБАЛОВИДНОЙ МЫШЦЫ КРЫСЫ В УСЛОВИЯХ ГРАВИТАЦИОННОЙ РАЗГРУЗКИ И ПОСЛЕДУЮЩЕГО ВОССТАНОВЛЕНИЯ
Автореферат кандидатской диссертации
На правах рукописи
МИРЗОЕВ Тимур Махмашарифович
ДИНАМИКА КЛЕТОЧНОГО ДЫХАНИЯ И СОДЕРЖАНИЕ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ В ВОЛОКНАХ КАМБАЛОВИДНОЙ МЫШЦЫ КРЫСЫ В УСЛОВИЯХ ГРАВИТАЦИОННОЙ РАЗГРУЗКИ И ПОСЛЕДУЮЩЕГО ВОССТАНОВЛЕНИЯ
03.03.01 - физиология
03.01.02 - биофизика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва - 2012
Работа выполнена в Федеральном государственном бюджетном учреждении науки Государственном научном центре Российской Федерации - Институте медико-биологических проблем Российской академии наук
НАУЧНЫЕ РУКОВОДИТЕЛИ:
доктор биологических наук, профессораа ааШенкман Борис Стивович
кандидат физико-математических наук Огнева Ирина Владимировна
ОФИЦИАЛЬНЫЕ ОППОНЕНТЫ:
доктор биологических наук, профессор,
заведующая лабораторией физиологии
мышечной деятельности
ГН - РФ - ИМБП РАНа Виноградова Ольга Леонидовна
доктор биологических наук, профессор,
заведующий научно-исследовательской
лабораторией адаптационной медицины
факультета фундаментальной медицины
ФГБОУ ВПО МГУ им. М.В. Ломоносов Архипенко Юрий Владимирович
аа
ВЕДУЩАЯ ОРГАНИЗАЦИЯ:
Институт биохимической физики имени Н.М. Эмануэля Российской академии наук
Защита диссертации состоится л28 _июня 2012 г. в 10-00 часов на заседании диссертационного совета Д 002.111.01 в Федеральном государственном бюджетном учреждении науки Государственном научном центре Российской Федерации Ч Институте медико-биологических проблем РАН по адресу: 123007, г. Москва, Хорошевское шоссе, д.76а.
С диссертацией можно ознакомиться в библиотеке Федерального государственного бюджетного учреждения науки Государственном научном центре Российской Федерации Ч Институте медико-биологических проблем РАН.
Автореферат разослан л25мая 2012 г.
Ученый секретарь диссертационного совета,
доктор биологических наук ааМ.А. Левинских
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Следствием длительного пребывания в условиях невесомости и гипокинезии является снижение функциональных возможностей мышечной системы и, в первую очередь, постуральных мышц, в частности камбаловидной. Именно камбаловидная мышца (m. soleus) несет основную нагрузку в поддержании вертикальной позы и противодействии земной гравитации, и, следовательно, при гравитационной разгрузке изменения в ней наиболее выражены (Fitts R. et al., 2000). Показано, что в условиях реальной или моделируемой гравитационной разгрузки имеет место развитие атрофических изменений в волокнах скелетных мышц млекопитающих и человека (Козловская И.Б. и др., 1984). Так, происходит снижение сократительных характеристик мышечных волокон (МВ) и их жёсткости (McDonald K.S., Fitts R.H., 1995; Toursel T. et al., 2002; Widrick J. et al., 1999; Ogneva I.V., 2010), отмечается уменьшение площади поперечного сечения мышечных волокон (Оганов В.С. и др., 1982; Riley D. et al., 2002) и трансформация миозинового фенотипа в быструю сторону (Caiozzo V. et al., 1994; Edgerton V. et al., 1995). Кроме того, наблюдаются изменения содержания сократительных белков (Chopard A. et al., 2001) и объема миофибриллярного аппарата (Kozlovskaya I.B. et al., 1996; Desplanches D. et al., 1987). Данные изменения приводят к снижению работоспособности постуральных мышц, что значительно осложняет осуществление длительных космических полётов и течение реабилитационного периодаа после их завершения.
Ряд последствий микрогравитации, имеющих место ва камбаловидной мышце, может быть связан с изменениями энергетического обмена, существенным аспектом которого является клеточное дыхание. Интенсивность клеточного дыхания способна оказывать влияние на параметры внутриклеточного энергообмена, в частности, ана динамику содержания таких энергетических субстратов, как гликоген и триглицериды (Baldwin K. et al.,1993; Widrick J. et al., 1999;а Grichko V. et al., 2000; Тавитова М.Г. и др., 2011).
Изменение уровня запасов внутриклеточных субстратов в условиях микрогравитации может быть связано как с трансформацией процесса клеточного дыхания, который зависита ота активности ферментов цикла Кребса, состояния цепи переноса электронов и проницаемости мембраны митохондрий для АДФ, так и с модуляцией энергопотребления в условиях сниженной сократительной активности мышцы. Литературные данные, свидетельствующие об активности ферментов цикла Кребса в условиях микрогравитации, являются противоречивыми (Буравкова Л.Б., Маилян Э.С., 1988;а Chi M.M. et al, 1992; Ohira Y. et al., 1992; Bigard A. et al., 1998; Shenkman B.S. et al., 2000). Однако о влиянии активности ферментов цикла Кребса на процесс клеточного дыхания можно говорить только в том случае, когда состояние дыхательной цепи остаётся интактным. Имеются данные о снижении активности цитохромоксидазы в волокнах камбаловидной мышцы крысы (Ohira Y. et al., 1994; Oishi Y. et al., 2008).а Кроме того, клеточное дыхание зависит от проницаемости митохондриальной мембраны, в регуляции которой, по-видимому, важную роль играют цитоскелетные белки (Saks V. et al., 1995). Возможно, одним из таких белков являетсяа десмин, что было показано в опытах с нуль-десминовыми мышами (Kay L. et. el., 1997; Milner D. et al., 2000). Было обнаружено, что у нокаутных по гену десмина мышейа интенсивность поглощения кислорода волокнами камбаловидной мышцы и константа диссоциации АДФ достоверно снижены по сравнению с теми же показателями у обычных мышей (Milner D. et al., 2000).а Также известно, что митохондрии находятся в тесной взаимосвязи с филаментами цитоскелетного белка десмина, который определяет их локализацию в клеткеа (Georgatos S.D., Maison C., 1996; Milner D. et al., 2000; Capetanaki Y. et al., 2007) и, возможно, участвует в регуляции транспорта АДФ и креатина через наружную мембрану митохондрий (Saks V. et al., 1995). Вследствие того, что десмин оказывает влияние на распределение и функции митохондрий, можно предположить, что изменения в содержании данного белка в условиях гравитационной разгрузки и после восстановления могут коррелировать с динамикой параметров клеточного дыхания.
С другой стороны, помимо митохондрий, филаменты десмина тесно связаны с Z-диском, одним из основных структурных белков которого является альфа-актинин-2 (Sanger J.M., Sanger J.W., 2008). Поэтому можно предположить, что содержание альфа-актинина-2 в МВ косвенно влияет на работу митохондрий, а, следовательно, и на процесс клеточного дыхания.а В то же время, на ранних этапах гравитационной разгрузки происходит снижение содержания десмина (Enns D.L. et al., 2007; Ogneva I.V., 2010), связанное, по-видимому, с активацией в этих условиях кальций-зависимых протеаз - кальпаинов (Enns D.L. et al., 2007), что и обусловливает деградацию белка. При этом субстратом кальпаиновой системы протеолиза является не только десмин, но и альфа-актинин-2 (Akiyama N. et al., 2006).
В настоящее время особенности клеточного дыханияа и факторы, влияющие на них, в мышечных волокнах в условиях микрогравитации и последующего восстановления изучены недостаточно. Следовательно, исследование параметров клеточного дыхания во взаимосвязи с содержанием цитоскелетныха белков и энергосубстратов в волокнах постуральных мышц является актуальным направлением в области современной гравитационной физиологии и клеточной биофизики.
В связи с вышеизложенным, целью работы являлось определение динамики клеточного дыхания во взаимосвязи с содержанием цитоскелетных белков и энергетических субстратов в волокнах камбаловидной мышцы крысы в условиях гравитационной разгрузки и последующего восстановления.
аЗадачи работы:
1.Определить основные параметры клеточного дыхания в волокнах камбаловидной мышцы как в динамике гравитационной разгрузки, так и последующего периода реадаптации.
2.Оценить динамику содержания десмина в мышечных волокнах камбаловидной мышцы в условиях гравитационной разгрузки и после восстановления.
3.Изучить изменение содержания альфа-актинина-2 в волокнах камбаловидной мышцы в условиях гравитационной разгрузки и после восстановления.
4.Определить содержание гликогена и триглицеридов в быстрых и медленных волокнах камбаловидной мышцы крысы в период 3 и 7- суточной реадаптации после 14-суточной гравитационной разгрузки.
Научная новизна работы:
-показаноа снижение параметров клеточного дыхания скинированных волокон камбаловидной мышцы крысы на ранних сроках гравитационной разгрузки, а также послеа 3-суточного восстановления, при этом послеа 7 суток реадаптации произошло восстановление скорости клеточного дыхания;
-показано снижение относительного содержания десмина после 3-суточого периода восстановления, а также восстановление данного белка к 7 суткам периода реадаптации;
-показано снижение содержания альфа-актинина-2 в волокнах камбаловидной мышцы крысы после 3,7,14 суток гравитационной разгрузки, а также 3-суточного восстановления; восстановление содержания альфа-актинина-2 до контрольного уровня после 7 суток реадаптации;
-показано увеличение содержания гликогена в быстрых волокнах на 3-и сутки реадаптации после гравитационной разгрузки, накопление гликогена в быстрых и медленных волокнах после 7-суточного восстановления;
-показано снижение содержания триглицеридов после 3-суточного восстановления в быстрых волокнах относительно показателей контрольной группы и в медленных волокнах относительно показателей в группе животных после вывешивания.
Научная и практическая значимость работы
Результаты, полученные в рамках данной работы, расширяют научные представления об особенностях процесса клеточного дыхания как в условиях гравитационной разгрузки, так и в последующий период реадаптации. В ходе работы были выявлены возможные факторы и механизмы, влияющие наа динамику параметров клеточного дыхания, а также содержание цитоскелетных белков и энергетических субстратов. В настоящем исследованииа была дана оценка изменениям, происходящим в камбаловидной мышце не только в условиях гравитационной разгрузки, но и в период реадаптации, что особенно важно для пониманияа процессов, имеющих место при реабилитации космонавтов после приземления на Землю.
Полученные в данной работе результаты могут быть использованы для разработки новых методов, которые позволят снизить нежелательное действие невесомости на постуральные мышцы космонавтов, совершающих длительные космические полёты.
Основные положения, выносимые на защиту:
1. Моделируемая гравитационная разгрузка приводит к достоверному снижению скорости клеточного дыхания волокон камбаловидной мышцы крысы, которая достигает минимума после 7 суток разгрузки, при этом наблюдается изменение содержания цитоскелетных белков (десмина и альфа-актинина-2).
2. Результатома восстановления в течение 3 суток является снижение скорости клеточного дыхания волокон камбаловидной мышцы. Реадаптация в течение 7 суток приводит ка восстановлению параметров клеточного дыхания. В период восстановления в волокнах камбаловидной мышцы отмечается изменение содержания цитоскелетных белков (десмина и альфа-актинина-2) и энергетических субстратов (гликогена и триглицеридов).
3. В условиях гравитационной разгрузки и в последующий период реадаптации интенсивность клеточного дыхания коррелирует с относительным содержанием десмина.
Публикации. Результаты диссертации изложены в 12 публикациях в научных журналах и сборниках тезисов конференций, из них 3 в журналах, рекомендованных ВАК.
Апробация работы. Основные результаты работы были доложены на VIа Всероссийской с международным участием школе-конференции по физиологии мышц и мышечной деятельности Системные и клеточные механизмы в физиологии двигательной системы (Москва, 2011); на XVIII Международной конференции студентов, аспирантов и молодых ученых Ломоносов-2011 (Москва, 2011); на X и XI Конференциях молодых учёных, специалистов и студентов, посвящённых Дню космонавтики (Москва, 2011, 2012); на XV Международной Пущинской школе-конференции молодых ученых Биология - наука XXI века (Пущино, 2011); на 40th European Muscle Conference (Берлин, Германия, 2011); на Космическом форуме 2011, посвящённом 50-летию полёта в космос Ю.А. Гагарина (Москва, 2011), на XIV международном совещании и VII школе по эволюционной физиологии (Санкт-Петербург, 2011).
Диссертация апробирована на заседании секции учёного совета ГН - РФ - ИМБП РАН Космическая физиология и биология (протокол № 2 от 06.03.2012г.).
Работа выполнена при поддержке программы фундаментальных исследований ГН - РФ - ИМБП РАН и гранта РФФИ 10-04-00106-а.
Структура диссертации. Работа включает в себя введение, обзор литературы, описание материалов и методов исследования, изложение результатов и их обсуждение, а также выводы. Диссертационная работа изложена на 114 страницах, содержит 32 рисунка, 6 таблиц и список цитируемой литературы из 188 источников.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Экспериментальные группы животных. В экспериментах использовали самцов крыс породы Вистара массой 180-220г, выращенных в питомнике ГН - РФ-ИМБП РАН.
В ходе работы были проведены 2 серии экспериментов. В первой серии экспериментов крысы были разделены на 5 групп, в каждой из которых было по семь животных: 1. Контроль -а группа животных, находившихся в стандартных виварных условиях при комнатной температуре около 23?С, астандартный комбинированный корм и воду ad libitum.а 2. л1-HS (HS - от англ. hindlimb suspension) - группа антиортостатического вывешивания крыс в течение одних суток. 3.а л3-HS - группа 3-суточного вывешивания. 4.а л7-HS - группа 7-суточного вывешивания. 5. л14-HS - группа 14-суточного вывешивания.
Во второй серии экспериментов крысы были разделены на 4 группы, в каждой из которых было по семь животных: 1. Контроль -а группа животных, находившихся в стандартных виварных условиях. 2. л14-HS - группа 14-суточного вывешивания. а3. л14-HS+3R (R - от англ. reloading) - группа 14-суточного вывешивания с последующим 3-х суточным восстановлением (в период восстановления крысы содержались в стандартных виварных условиях, как и контрольная группа). 4. л14-HS+7R - группа 14-суточного вывешивания с последующим восстановлением в течение 7 суток. Все процедуры с животными были одобрены комиссией по биомедицинской этике ГН - РФ-ИМБП РАН.
Метод антиортостатического вывешивания крыс. Для моделирования гравитационной разгрузки применялся метод антиортостатического вывешивания по Ильину-Новикову в модификации Морей-Холтон (Morey-Holton E.R., Globus R.K., 2002). Животных подвешивали на подвижной цепочке за хвост таким образом, чтобы угол между осью тела и поверхностью земли составлял 30. Вывешенным животным были доступны вода и корм adlibitum.
Материал исследования. аМатериалом исследования служила камбаловидная мышца - m. soleus, которую выделяли из задних конечностей крыс. Для взятия биоматериала крысы наркотизировались раствором 10% авертина в концентрации 4 мл/кг веса тела животного. Сразу же после взятия биоматериала животные умерщвлялись введением повышенной дозы авертина (10 мл/кг).
Для иммуногистохимического окрашивания на субстраты и выделения белка, полученные кусочки ткани ориентировали поперечно, затем фиксировали на бумажной подложке с помощью Tissue-Tekо (OCTTM Compound 4583) и замораживали в жидком азоте. Мышечные пробы хранили при температуре -80С вплоть до обработки. Для дальнейшей иммуногистохимической окраски в криостате фирмы Leica при -20Са делали поперечные срезы мышечной ткани толщиной 10 мкм. Срезы располагали на покрытых адгезивом гистологических стеклах. Другая часть замороженного материалаа нарезалась в пробирки с целью дальнейшего выделения белка.а
Для определения скорости клеточного дыхания кусочки мышечной ткани не подвергались заморозке, а были помещены в специальный раствор сразу после выделения из животного.
Метод полярографии. Исследуемую мышцу вырезали от сухожилия до сухожилия и немедленно помещали в охлажденный раствор А, в котором ее разделяли на пучки мышечных волокон длинной 3-4 мм и толщиной около 1 мм. Далее волокна инкубировали в растворе А с сапонином (50 мкг/мл) в течение 30 минут при легком помешивании, при температуре 40C с целью частичного скинирования мембраны. Затем пучки волокон отмывали от сапонина в течение 10 минута в растворе Б. Скорость поглощения кислорода оценивали методом полярографии по Саксу (Saks V.A., et al., 1998). Пучки скинированных волокон инкубировали в растворе Б, в который добавляли 2 мг/мл бычьего сывороточного альбумина (БСA), свободного от жирных кислот. Измерение концентрации кислорода проводилиа с помощью электрода Кларка и оксиметра YSIа model 53 oxygen monitorа (Yellow Spring Instrument Co., USA) при температуре 220C. Растворимость кислорода в 1 мл среды инкубации при этой температуре принимали равной 460 нг-ат (Seppet E.K. et al., 2004).
Для оценки клеточного дыхания были выбраны следующие параметры: Vбаз. - скорость потребления кислорода без добавления в среду субстратов. VМ/Gl - скорость потребления кислорода на экзогенных субстратах, в качестве которых использовались 2 мМ малат и 5 мМ глутамат. Выбор данных субстратов дыхания был сделан на основании того, что данные субстраты поставляют электроны на первый комплекс дыхательной цепи, следовательно, это позволило судить о структурно-функциональной целостности системы переносчиков на внутренней мембране митохондрий. VАДФ - скорость потребления кислорода при добавлении АДФ (0,1 мМ), что позволило судить о состоянии систем транспорта нефосфорилированных макроэргов, а также состоянии терминального участка дыхательной цепи. После измерений волокна извлекали из полярографической ячейки, высушивали при температуре +950С и затем взвешивали, что позволяло рассчитывать скоростные параметры, нормированные на г сухого веса. Дыхательный контроль (ДК) рассчитывали как отношение скорости дыхания в присутствии АДФ к скорости дыхания на малате и глутамате.а
Метод гель-электрофореза с последующим иммуноблоттингом. Электрофорез по методу Лэммли (Остерман Л.А., 1981) проводили в 12% разделяющем полиакриламидном геле и 6% концентрирующем геле. Электрофорез проводили в мини-системе Bio-Rad (США) в течение часа при силе тока 20мА на один гель. С учётом проведённого измерения концентрации, в каждую лунку наносили по 5мкг белка.
Перенос белков на нитроцеллюлозную мембрану (Towbin H.et al., 1979) осуществлялиа в системе mini Trans-Blot (Bio-Rad) в течение 12 часов при температуре +4С и силе тока 150мА. После переноса мембрану блокировали в 4% растворе сухого молока в PBST (PBS + 0,1% Tween 20) в течение часа при комнатной температуре. Затем проводили инкубацию с первичными моноклональными антителами (Santa-Cruz, США), а также с биотинилированными вторичными козьими антителами против IgG мышей (Sigma, США) в разведении. Дляа визуализации полос использовали перекись водорода с 3,3Т- диаминобензидином (Merck, Германия).а Затем мембраны сканировали и анализировали с использованием компьютерной программы ImageJ, определяя содержание белка по оптической плотности относительно контроля.а
Гистохимическое окрашивание на гликоген с одновременным типированием мышечных волокон. Для аопределения содержания гликогена в быстрых и медленных МВ камбаловидной мышцы использовали йодную кислоту, реактив Шиффа, а также первичные и вторичные антитела против быстрых и медленных изоформ тяжёлых цепей миозина (Schaart G. et al., 2004; Тавитова М.Г и соавт., 2011).
Гистохимическое окрашивание на триглицериды с одновременным типированием мышечных волокон. Для определения содержанияа триглицеридов в быстрых и медленных волокнах использовали масляный красный (Oil red O) в сочетании с первичными и вторичными антителами. (Koopman et al., 2001; Тавитова М.Г. и соавт., 2011).
Анализ окрашенных срезов проводили с помощью флуоресцентного микроскопа фирмы Leica (Германия), снабжённого светофильтрами, видеокамерой Leica DC 300F, а также системой компьютерного анализа изображения. Все препараты мышечных срезов фотографировали при 20-кратном увеличении. Полученные фотографии обрабатывали в специальной программе Leica QWin. Для определения содержания триглицеридов один и тот же срез фотографировали сначала под зелёным фильтром, а затем под синим фильтром. В зелёном спектре выявляли результаты окраски на триглицериды (красная флуоресценция разной интенсивности). В синем спектре определяли волокна, содержащие различные изоформы тяжёлых цепей миозина, характеризующиеся зелёным свечением. Затем, сопоставляя оба изображения, измеряли интенсивность флуоресценции, свидетельствующую о содержании триглицеридов в быстрых и медленных мышечных волокнах. Для определения содержания гликогена один и тот же срез мышечной ткани фотографировали сначала в видимом свете, а затем под синим фильтром. Сравнивая полученные изображения, измеряли содержание гликогена по оптической плотности. Значения содержания триглицеридов и гликогена выражали в условных единицах.
Статистическая обработка. Оценку значимости различия средних значений показателя в независимых выборках проводили по t-критерию Стьюдента с помощью стандартной программы Microsoft Excel 2007о. Результаты представляли в виде Мm, где М - среднее арифметическое значение показателя в сравниваемых группах, m - ошибка среднего значения. Различие средних значений показателя в сравниваемых выборках считали достоверными при уровне значимости p?0,05.
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТОВ И ИХ ОБСУЖДЕНИЕ
Динамика скорости клеточного дыхания и содержание цитоскелетных белков в период гравитационной разгрузки. Полученные результаты свидетельствуют о том, что интенсивность клеточного дыхания снизилась через трое суток гравитационной разгрузки, достигла минимума через семь суток и несколько возросла к четырнадцати суткам моделируемой разгрузки (табл.1).
Таблица 1.
Параметры клеточного дыхания в волокнах камбаловидной мышцы крысы на разных сроках гравитационной разгрузки.
Параметр Группа |
Vбаз, нмоль О/минг |
VМ/Gl, нмоль О/минг |
VАДФ, нмоль О/минг |
ДК |
Контроль |
1082166 |
139098 |
1698158 |
1,140.19 |
1-HS |
95084 |
1306158 |
157282 |
1,280.11 |
3-HS |
762130* |
1074114* |
153486* |
1,680.13* |
7-HS |
620110* |
756114* |
868130* |
1,330.28 |
14-HS |
736110* |
110686*/& |
1374172*/& |
1,510.29 |
*- достоверное отличие от группы Контроль (p<0,05), & - достоверное отличие от группы л7-HS (p<0,05).
Отмечавшиеся в данном исследовании изменения параметров клеточного дыхания в условиях функциональной разгрузки могли быть связаны с активностью ферментов цикла Кребса, состоянием комплексов дыхательной ацепи, а также проницаемостью митохондриальной мембраны для макроэргов (АДФ и креатина). Рядом авторов было отмечено, что воздействие 2-3-х недельнойаа гравитационной разгрузки на камбаловидную мышцуа не приводит к достоверным изменениям активности малатдегидрогеназы (Chi M. et al., 1992), сукцинатдегидрогеназы (Ohira Y. et al., 1992; Shenkman B.S. et al., 2000) и цитратсинтазы (Bigard A. et al., 1998). Однако Ohira Y. et al. (1994) показали снижениеа активности цитратсинтазы в камбаловидной мышце после 10-суточного вывешивания крыс. О влиянии активности ферментов цикла Кребса на процесс клеточного дыхания можно говорить только в том случае, когда состояние дыхательной цепи остаётся неизменным. В работах Ohira Y. et al. (1994) и Oishi Y. et al. (2008) было показано, что гравитационная разгрузка в течение 10 и 14 суток приводила к снижению активности цитохромоксидазы в волокнах камбаловидной мышцы крысы. Данный факт может свидетельствовать о нарушении работы дыхательной цепи митохондрий, так как цитохромоксидаза катализирует конечный этап переноса электронов на кислород в процессе окислительного фосфорилирования. Снижение скорости клеточного дыхания мышечных волокон в условиях разгрузки могло быть связано с изменением активности 5Т-аденозинмонофосфат-зависимой протеинкиназы (AMPK). Winder W. W. et al., (2000) было показано, что при активации AMPK в МВ I типа m. soleus крысы произошло достоверное увеличение активности ряда митохондриальных ферментов: цитратсинтазы, малатдегидрогеназы, сукцинатдегидрогеназы, а также цитохрома с. Группой учёных Han B. et al. (2007) показано, что гравитационная разгрузка в течение двух недель привела к достоверному снижению активности AMPK в m. soleus крысы. Известно, что АМФ-зависимая протеинкиназа ингибируется АТФ и креатинфосфатом, а активируется повышенной концентрацией АМФ (Corton J.M. et al., 1994; Hawley S.A. et al., 1996). Накопление фосфорилированных форм макроэргических фосфатов в мышечных волокнах при гравитационной разгрузке (Wakatsuki T. et al., 1994) могло обусловливать снижение активности 5Т-аденозинмонофосфат-зависимой протеинкиназы (Han B. et al., 2007), осуществляющей контроль над экспрессией митохондриальных белков (Winder W.W., 2001). Всё это могло привести к снижению концентрации комплексов дыхательной цепи в митохондриях и уменьшению V М/Gl и VАДФ, которые по нашим данным были минимальны через семь суток гравитационной разгрузки.
После 14 суток антиортостатического вывешивания скорость дыхания несколько возросла относительно показателейа 7-суточной разгрузки. Возможно, это могло бытьа следствием увеличения активных комплексов дыхательной цепи митохондрий. В то же время Bigard A.X. et al. (1998) было показано, что после 3-х недель антиортостатического вывешивания потребление кислорода волокнами камбаловидной мышцы относительно контрольных значений не менялось. В работе Nemirovskaya T.L., Shenkman B.S. (2002) было отмечено увеличение максимальной скорости дыхания волокон камбаловидной мышцы после двухнедельного вывешивания. По полученным в настоящем исследованииа данным, в период с 7 по 14 сутки разгрузки происходит достоверный рост параметров клеточного дыхания. Поэтому, можно предположить, что при более длительных сроках вывешивания скорость клеточного дыхания могла возрасти до уровня контроля или даже превысить его в связи с тем, что содержание десмина после двух недель разгрузки восстанавливается. аРост скорости потребления кислорода волокнами камбаловидной мышцы с 7 по 14 сутки разгрузки соотносится с данными Alford E.K. et al. (1987), показавшими увеличение электромиографической активности m. soleus в данный период.
Известно, что белок цитоскелета десмин тесно связан с митохондриями и определяет их локализацию и функции (Milner D. et al., 2000; Capetanaki Y. et al., 2007). аБыло показано, что у нуль-десминовых мышей происходит нарушение нормального распределения и локализации митохондрий в волокнах камбаловидной мышцы, что сопровождаетсяа снижением интенсивностиа клеточного дыхания (VАДФ) (Kay L. et al., 1997; Milner D. et al. 2000). Кроме того, разрушение десмина могло привести к нарушению проницаемости внешней митохондриальной мембраны для АДФ и креатина (Saks V.A. et al., 1995). Следует также отметить, что на состояние десмина может оказывать влияние структурный белок Z-диска альфа-актинин-2, так как десмин тесно связан с Z-диском саркомера (Sanger J.M., Sanger J.W., 2008). Таким образом, полученные результаты параметров клеточного дыхания волокон камбаловидной мышцы в динамике гравитационной разгрузки и в период реадаптации могли быть вызваны изменениями работы митохондрий, на функции которых, по-видимому, влияют цитоскелетные белки (Saks V.A. et al., 1995). Поэтому одной из целей данной работы являлось определение относительного содержания таких цитоскелетных белков, как альфа-актинин-2 и десмин.
Полученные в данном исследовании результаты по содержанию десмина в волокнах камбаловидной мышцы в процессе разгрузки (табл.2) согласуются с данными, описанными ранее в работе Enns D.L. et al. (2007) и Ogneva I.V. (2010). Так, в исследовании Ogneva I.V. (2010) было показано достоверное снижение относительного содержания десмина после 3-суточного вывешивания, максимальное снижение после 7 суток разгрузки и восстановление содержания десмина после 12 суток вывешивания.а Кроме того, Enns D.L. et al. (2007) показали, что после гравитационной разгрузки содержание десмина в широкой мышце бедра крысы снижается уже к первым суткам разгрузки, остаётся ниже контроля после 3-х суток и практически полностью восстанавливается к девятым суткам. Десмин и альфа-актинин-2 являются субстратами кальций-зависимой системы протеолиза - кальпаиновой системы (Goll D.E. et al., 2003) увеличение активности работы которой в условиях гравитационной разгрузки показано Enns D.L., Belcastro A.N. (2006), Enns D.L. et al., (2007), Алтаевой Э.Г. и соавт., (2010), Ma X.W et al. (2011). Наблюдавшееся восстановление содержания десмина в камбаловидной мышце к 14 суткам разгрузки согласуется са данными Chopard A. et al. (2001), показавшими, чтоа при длительных сроках вывешивания (3 и 6 недель) достоверных отличий от уровня контроля в относительном содержании десмина в камбаловидной мышце крысы не наблюдалось. В то же время, содержание альфа-актинина-2 после двухнедельного вывешивания восстановлено не было (табл. 3), что могло свидетельствовать о большей его деградации и /или оа более замедленном компенсаторном механизме, активирующим синтез данного белка
Таблица 2.
Содержание десмина в волокнах камбаловидной мышцы крысы на разных сроках гравитационной разгрузки
Группа |
Относительное содержание десмина, % от контроля |
Контроль |
1005 |
1-HS |
1047 |
3-HS |
806* |
7-HS |
673* |
14-HS |
924& |
*- достоверное отличие от группы Контроль (p<0,05),а & - достоверное отличие от группы л7-HS (p<0,05).
Таблица 3.
Содержание альфа-актинина-2 в волокнах камбаловидной мышцы крысы на
разных сроках гравитационной разгрузки
Группа |
Относительное содержание альфа-актинина-2, % от контроля |
Контроль |
1007 |
1-HS |
1028 |
3-HS |
866* |
7-HS |
748* |
14-HS |
866*/& |
а а* - достоверное отличие от группы Контроль (p<0,05),а
& - достоверное отличие от группы л7-HS (p<0,05).
а Динамика скорости клеточного дыхания и содержание цитоскелетных белков в период восстановления после гравитационной разгрузки. После 3-х суток восстановления было обнаружено снижение исследуемых параметров клеточного дыхания как относительно контроля, так и относительно 14-суточного вывешивания (табл.4). Подобное снижение скорости клеточного дыхания в период реадаптации было показано Маилян Э.С. и Буравковой Л.Б. (1983) у крыс, подвергавшихся невесомости в течение 18,5 суток на биоспутнике Космос-1129. Однако исследование проводилось на суспензии митохондрий, выделенных из задней группы мышц бедра, состоящих преимущественно из быстрых волокон. После 7 суток реадаптации было обнаружено восстановление всех параметров скорости клеточного дыхания почти до контрольного уровня, что могло свидетельствовать о восстановлении функциональных способностей митохондрий в волокнах камбаловидной мышцы.
Таблица 4.
Параметры клеточного дыхания в волокнах камбаловидной мышцы крысы в период восстановления после гравитационной разгрузки.
Параметр Группа |
Vбаз, нмоль О/минг |
VМ/Gl, нмоль О/минг |
VАДФ, нмоль О/минг |
ДК |
Контроль |
1082166 |
139098 |
1698158 |
1,140.19 |
14-HS |
736110* |
110686* |
1374172* |
1,510.29 |
14-HS + 3R |
49488*/$ |
640104*/$ |
952106*/$ |
1,480.28 |
14-HS + 7R |
1040156$ |
1276136$ |
1624112$ |
1,370.27 |
а*- достоверное отличие от группы Контроль (p<0,05), $ - достоверное отличие от группы л14-HS (p<0,05).
После 3-суточного периода восстановления произошло снижение относительного содержания десмина относительно группы лконтроль и л14-HS и альфа-актинина-2 относительно группы лконтроль. Период реадаптации в течение 7 суток привёл к восстановлению содержания исследуемых белков (табл.5, 6).а
Снижение содержания десмина и альфа-актинина-2 в острый период реадаптации коррелирует с электронномикроскопическими данными Riley D.A. et al. (1987), которые показали, что ранний период реадаптации атрофированной мышцей воспринимается как эксцентрическая нагрузка, поскольку повреждения структуры саркомера, наблюдаемые через 4,5 часа восстановления аналогичны таковым после эксцентрических сокращений (Riley D.A. et al., 1990, 1992, 2000). Резкая потеря десмина волокнами скелетных мышц, подвергавшихся эксцентрическойа нагрузке, была показана в работах Barash I. et al. (2002), Lieber R. et al. (1996), Friden J., Lieber R. (1998). Friden J. и Lieber R. (1996) были обнаружены серьёзные нарушения структуры Z-диска саркомера в волокнах скелетных мышц кролика после эксцентрической нагрузки. Следовательно, резкое снижение скорости клеточного дыхания после 3-х суток реадаптации могло быть вызвано нарушением работы митохондрий вследствие деградации десмина и альфа-актинина-2.
Таблица 5.
Содержание десмина в волокнах камбаловидной мышцы крысы в период восстановления после гравитационной разгрузки
Группа |
Относительное содержание десмина, % от контроля |
Контроль |
1005 |
14-HS |
924 |
14HS+3R |
аа 843*/$ |
14HS+7R |
1004 |
*- достоверное отличие от группы Контроль (p<0,05), $ - достоверное отличие от группы л14-HS (p<0,05).
Таблица 6.
Содержание альфа-актинина-2 в волокнах камбаловидной мышцы крысы в период восстановления после гравитационной разгрузки
Группа |
Относительное содержание альфа-актинина-2, % от контроля |
Контроль |
1007 |
14-HS |
866* |
14-HS+3R |
873* |
14-HS+7R |
1029 |
аа
* - достоверное отличие от группы Контроль (p<0,05).
В ходе работы был проведён корреляционный анализ динамики параметров клеточного дыхания и содержания десмина при гравитационной разгрузке и в период реадаптации. Данный анализ выявил наличиеа выраженной корреляции между изменениями содержания десмина и скоростью клеточного дыхания (VАДФ), при этом коэффициент ранговой корреляции аСпирмена (rs) составил а0,78.
Содержание триглицеридов в медленных и быстрых МВ камбаловидной мышцы в период реадаптации. Увеличение содержания триглицеридов в медленных МВ (рис.1) могло быть связано с более низким расходованием данного субстрата в связи с низким энергопотреблением неактивной мышцей и, соответственно, низким окислением жирных кислот в митохондриях. Это согласуется с данными Baldwin K.M. et al. (1993) о снижении способности промежуточной широкой мышцы бедра окислять жирные кислоты после 9-суточного полёта. Антиортостатическое вывешивание не привело к достоверным изменениям содержания триглицеридов в быстрых мышечных волокнах (рис.2). Вероятно, это связано с тем, что для данных волокон характерен гликолитический тип окисления, для которогоа главным субстратом является гликоген, а не триглицериды. Отсутствие достоверных отличий от контроля по содержанию триглицеридов в широкой мышце бедра (состоящей преимущественно из МВ быстрого типа) после реальной и модулируемой микрогравитации было отмечено Musacchia X.J et al., (1992).
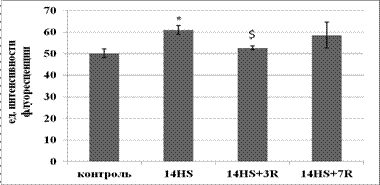
Рис.1. Содержание триглицеридов в медленных МВ камбаловидной мышцы.
* - достоверное отличие от группы лконтроль (p<0,05),$ - достоверное отличие от группы л14-HS (p<0,05).
Период реадаптации в течение 3-х суток обнаружил достоверное снижение содержания триглицеридов в быстрых МВ относительно контроля и в медленных МВ относительно группы вывешивания. Возможно, данные изменения связаны с тем, что ранний период реадаптации воспринимался мышцей как эксцентрическая нагрузка (Riley D.A. et al., 1996). В связи с этим на клеточном уровне могли произойти изменения, активирующие сигнальные пути (возможно через активацию AMPK), приводящие к увеличению окисления жирных кислот. После 7 суток периода реадаптации произошло восстановление содержания триглицеридов до уровня контроля, что могло свидетельствовать оа нормализации метаболизма жиров в быстрых и медленных мышечных волокнах камбаловидной мышцы.
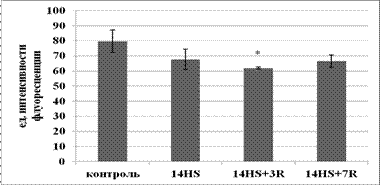
Рис. 2. Содержание триглицеридов в быстрых МВ камбаловидной мышцы.
* - достоверное отличие от группы лконтроль (p<0,05).
Содержание гликогена в медленных и быстрых МВ камбаловидной мышцы в период реадаптации. В медленных мышечных волокнах (рис.3) после разгрузки отличий от контроля не наблюдалось. Эти данные согласуются с результатами Тавитовой М.Г. и соавторов (2011), согласно которым после 14-суточного вывешивания содержание гликогена в волокнах камбаловидной мышцы крысы не отличалось от контроля. Возможно, это обусловлено тем, что медленные волокна, являясь по характеру метаболизма окислительными, меньше зависят от процессов, связанных с обменом гликогена. В мышечных волокнах быстрого типа (рис.4) произошло незначительное снижение содержания гликогена. Это могло быть связано с изменением уровня нервной активации. Так, согласно Alford E.K. et al. (1987) электромиографическая аактивность камбаловидной мышцы крысы уже через двенадцать суток вывешивания не отличалась от контроля.
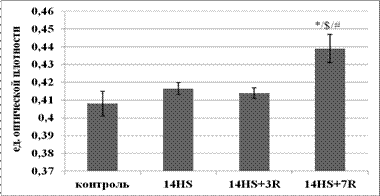
Рис. 3. Содержание гликогена в медленных МВ камбаловидной мышцы.
* - достоверное отличие от группы лконтроль (p<0,05),а $ - достоверное отличие от группы л14HS (p<0,05),а # - достоверное отличие от группы л14HS+3R (p<0,05).
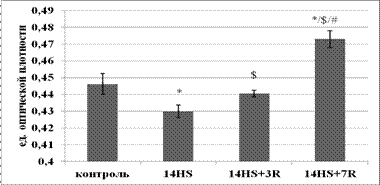
Рис. 4. Содержание гликогена в быстрых МВ камбаловидной мышцы.
* - достоверное отличие от группы лконтроль (p<0,05),а $ - достоверное отличие от группы л14HS (p<0,05),а # - достоверное отличие от группы л14HS+3R (p<0,05).
В быстрых волокнах после 3-х суток восстановления произошло увеличение количества гликогена по сравнению с группой вывешивания. Согласно данным Henriksen E. et al. (1988) в период восстановления после гравитационной разгрузки в камбаловидной мышце крысы произошло увеличение потребления 2-деоксиглюкозы по сравнению с уровнем контроля. Можно предположить, что повышение содержания глюкозы активировало синтез гликогена, что и привело к его накоплению после 3-х суток реадаптации. Накопление гликогена в быстрых и медленных МВ после 7-суточного восстановления могло быть связано с AMPK-сигнальной системой. Так, Merrill G. F. et al. (1997) было показано, что AMPK стимулирует не только окисление жирных кислот, но и увеличивает поступление глюкозы в клетку.
Следовательно, содержание энергетических субстратов в волокнах камбаловиднойа мышцы в ходе гравитационной разгрузки и последующей реадаптации подверглись изменениям, которые, по-видимому, явились следствием изменения внутриклеточного энергообмена.
Таким образом, в рамках данной работы была отслежена динамика параметров клеточного дыхания волокон камбаловидной мышцы во взаимосвязи с содержанием цитоскелетных белков как в условиях гравитационной разгрузки, так и в период реадаптации. При этом была установлена акорреляция между асодержанием цитоскелетного белка десмина и интенсивностью поглощения кислорода волокнами камбаловидной мышцы, что могло свидетельствовать о важной роли данного белка в регуляции клеточного дыхания. Кроме того, было установлено изменение содержания энергетических субстратов в волокнах камбаловидной мышцы в период реадаптации после гравитационной разгрузки.
ВЫВОДЫ
1. В условиях функциональной разгрузки задних конечностей крысы и последующий период восстановления в волокнах камбаловидной мышцы установлена тесная взаимосвязь между содержанием десмина и скоростью клеточного дыхания (VАДФ).
2. После 3, 7 и 14 суток функциональной разгрузки задних конечностей крысы произошло снижение скорости клеточного дыхания (Vбаз, VM/Gl, VАДФ) волокон камбаловидной мышцы относительно показателей контрольной группы; после 3-суточного восстановления произошло снижение всех показателей скорости клеточного дыхания относительно контроля; после 7 суток реадаптации наблюдалось восстановление параметров клеточного дыхания.
3. После 3 и 7 суток вывешиванияа крыс было отмечено снижение содержания десмина в волокнах камбаловидной мышцы по сравнению с контролем, к 14 суткам разгрузки произошло восстановление содержания десмина; после 3 суток периода восстановления наблюдалось снижение содержания десмина относительно группы 14-суточного вывешивания, к 7 суткам реадаптации произошло восстановление содержания десмина до контрольного уровня.
4.Гравитационная разгрузка задних конечностей крысы привела к снижению уровня содержания альфа-актинина-2 в волокнах камбаловидной мышцы относительно контроля, а также после 3 суток восстановления; на 7 сутки реадаптации произошло восстановление содержания альфа-актинина-2 до контрольного уровня.
5. После 7 суток восстановления было отмечено повышение содержания гликогена в медленных и быстрых волокнах камбаловидной мышцы крысы относительно значений гравитационной разгрузкиа и 3-суточного восстановления. После 3 суток реадаптации наблюдалось снижение содержания триглицеридов относительно значений гравитационной разгрузки в медленных мышечных волокнах камбаловидной мышцы крысы.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ ДИССЕРТАЦИИ
аа Статьи в журналах, рекомендованных ВАК:
- Мирзоев Т.М., Бирюков Н.С., Веселова О.М., Ларина И.М., Шенкман Б.С., Огнева И.В. Содержание десмина и клеточное дыхание волокон камбаловидной мышцы крысы в условиях 3- и 7-суточного восстановления после 14-суточного антиортостатического вывешивания. // Авиакосмическая и экологическая медицина. - 2012. - Т. 46. №1. - С. 41-46.
- Мирзоев Т.М., Бирюков Н.С., Веселова О.М., Ларина И.М., Шенкман Б.С., Огнева И.В. Параметры клеточного дыхания волокон и содержание десмина в камбаловидной мышце крысы на ранних этапах гравитационной разгрузки. // Биофизика. - 2012. - Т. 57. № 3. - C. 509-514.
- Мирзоев Т.М., Шенкман Б.С., Ушаков И.Б., Огнева И.В. Содержание десмина и альфа-актинина-2 в камбаловидной мышце крысы в динамике гравитационной разгрузки и последующего периода восстановления. // Доклады Академии наук. - 2012. - Т. 444. № 2.- С. 216-218.
Тезисы конференций:
- Мирзоев Т.М., Огнева И.В. Содержание энергетических субстратов в волокнах камбаловидной мышцы молодых крыс после 14-суточного антиортостатического вывешивания и 3-суточного восстановления. // Системные и клеточные механизмы в физиологии двигательной системы. Материалыа VIа Всероссийской с международным участием школы-конференции по физиологии мышц и мышечной деятельности. Москва. - 2011. - С.а 29.
- Мирзоев Т.М. Относительное содержание альфа-актинина-2 в волокнах камбаловидной мышцы крысы при функциональной разгрузке и восстановлении. // Ломоносов - 2011: XVIII Международная конференция студентов, аспирантов и молодых ученых; секция Биология; 11-15 апреля 2011 г; Москва, МГУ имени М.В. Ломоносова, биологический факультет: Тезисы докладов. Москва. - 2011. - С. 264-265.
- Мирзоев Т.М. Влияние гравитационной разгрузки на содержание десмина и энергетических субстратов в волокнах камбаловидной мышцы крысы. // X Конференция молодых ученых, специалистов и студентов, посвященная Дню космонавтики. Материалы конференции. - Москва. - 2011. - С. 51.
- Мирзоев Т.М., Огнева И.В. Влияние функциональной разгрузки на содержание десмина и альфа-актинина-2 в волокнах камбаловидной мышцы крысы. // Биологияа - наука ХXI века: 15-я Международная Пущинская школа-конференция молодых ученых (Пущино, 18-22 апреля 2011 года). Сборник тезисов. - Пущино. - 2011. - С. 140 - 141.
- Mirzoev T., Ogneva I. Correlation between desmin content and cell respiration in rat soleus fibers at early stages of gravitational unloading and after reloading. // The 40th European Muscle Conference. - Berlin. - 2011. - P. 84.
- аBiryukov N., Mirzoev T., Ogneva I. Transversal stiffness of Z-disk and alpha-actinin-2 content in the rat soleus muscle after 3- and 7 day reloading after 14-day hindlimb suspension. // The 40th European Muscle Conference. - Berlin. - 2001. - P. 59.
- Мирзоев Т.М., Огнева И.В. Содержание десмина и параметры клеточного дыхания волокон камбаловидной мышцы крысы в период реадаптации после гравитационной разгрузки. // Космический форум 2011, посвящённый 50-летию полёта в космос Ю.А. Гагарина (сборник материалов). - Москва. - 2011. - С. 190.
- аМирзоев Т.М., Огнева И.В. Динамика изменения параметров клеточного дыхания и содержания десмина в волокнах камбаловидной мышцы крысы на разных сроках функциональной разгрузки. // Четырнадцатое международное совещание и седьмая школа по эволюционной физиологии. Тезисы докладов и лекций. Санкт-Петербург. - 2011. - С. 127-128.
- Мирзоев Т.М. Динамика параметров клеточного дыхания и содержание цитоскелетных белков в волокнах камбаловидной мышцы крысы в период восстановления после гравитационной разгрузки. // XI Конференция молодых ученых, специалистов и студентов, посвященная Дню космонавтики. Материалы конференции. - Москва. - 2012. - С. 36-37.
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]