
 Все авторефераты докторских диссертаций
Все авторефераты докторских диссертаций
Птицы горных систем юга Восточной Сибири: экологические аспекты формирования и динамики их населения
Автореферат докторской диссертации
На правах рукописи
Ананин
Александр Афанасьевич
Птицы горных систем юга Восточной Сибири:
экологические аспекты формирования и динамики их населения
Специальность 03.02.08 - экология (биологические науки)
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Улан-Удэ
2012
Работа выполнена в ФГБОУ ВПО Бурятский государственный университет
Научный консультант: аа доктор биологических наук, профессор
Доржиев Цыдыпжап Заятуевич
Официальные оппоненты:а доктор биологических наук, профессор, ФГБОУ ВПО Красноярский государственный педагогический университет им. В.П. Астафьева
Баранов Александр Алексеевич
доктор биологических наук, профессор,
ФГБОУ ВПО Восточно-Сибирская
государственная академия образования
Пыжьянов Сергей Владимирович
доктор биологических наук, в.н.с.,
Институт биологических проблем
криолитозоны СО РАН
Дегтярев Виктор Григорьевич
Ведущая организация:аа ФГБОУ ВПО Томский государственный
университет
Защита состоится 21 мая 2012 года в ___ часов на заседании диссертационного совета Д 212.022.03 по защите диссертаций на соискание ученой степени доктора наук при Бурятском государственном университете по адресу: 670000 г. Улан-Удэ, ул. Смолина, 24а, Конференц-зал.
Факс: (3012) 210588;а e-mail: d21202203@mail.ru
С диссертацией можно ознакомиться в библиотеке
Бурятского государственного университета
Автореферат разослана ____ апреля 2012 г.
Ученый секретарь диссертационного совет аа
кандидат биологических наук аа а Н.А. Шорноева
Общая характеристика работы
Актуальность проблемы. Исследования долговременных изменений численности популяций животных в природных условиях по-прежнему сохраняют свою актуальность. Особая значимость таких работ связана с современными тенденциями роста воздействия антропогенных факторов и глобального изменения климата.
Изучение механизмов формирования видового населения птиц в горных условиях и факторов, влияющих на эти процессы - одно из слабо разработанных направлений исследования долговременной динамики численности. Особую значимость выполнение таких работ приобретает в горных системах юга Восточной Сибири. Эти горы лежат в южной части тайги, на стыке таежной и лесостепной зон. Характерное отличие гор региона заключается в том, что здесь горные леса не представляют собой экологические изоляты, а смыкаются с таежными массивами равнин и предгорий Восточной Сибири.
Выполнение долговременных наблюдений за динамикой обилия птиц в Северной Азии, в том числе на юге Восточной Сибири, имеет особое значение в связи с тем, что зимовки перелетных птиц этого региона располагаются главным образом в Юго-Восточной Азии и Индии, в отличие от европейской территории с преимущественно африканскими зимовками дальних мигрантов. Поскольку условия южно-азиатских и африканских зимовок могут существенно различаться, наблюдения за характером долговременных изменений численности местных гнездящихся популяций способствуют получению ответа об относительной роли рождаемости (успешности размножения) или постнатальной смертности (в период миграций и зимовки) в долговременных колебаниях плотности популяций.
Длительные стационарные наблюдения за обилием вида позволяют выявить особенности его реагирования на те или иные изменения условий среды, которые складываются в конкретный год. Данные по динамике обилия видов птиц одного и того же участка местности на многолетних отрезках времени имеют самостоятельную ценность уже потому, что таких данных по разным регионам немного, в том числе и по долговременным исследованиям сообществ лесных птиц (Enemar et al., 1994). Такие исследования осуществлялись, как правило, на небольших площадках (10-40 га), часто расположенных в изолированных лесных массивах. На территории Сибири 15-летние наблюдения (1978-1993 гг.) на площади 450 га выполнены на Енисейской экологической станции ИПЭЭ РАН в северо-таежной зоне (Бурский, 1986, 1993, 2007) и 20-летние - на площадках свыше 10 км? (1990-2009 гг.) - в лесотундровой зоне на севере Западной Сибири (Головатин, 2009). Долговременные исследования динамики обилия птиц протяженностью свыше 25 лет на достаточно обширных стационарных участках в горных условиях ранее не осуществлялись.
Цели и задачи исследования. Цель настоящего исследования - выявление основных закономерностей формирования видового населения птиц и факторов, определяющих динамику этих процессов в горных условиях.
Основные задачи исследования:
1. Выявить особенности структуры фауны и населения птиц в условиях горных систем юга Восточной Сибири.
2. Установить межгодовые флуктуации и закономерности многолетней динамики обилия гнездящихся и зимующих птиц в горных условиях на примере Баргузинского хребта.
3. Дать оценку влияния районов и условий зимовок на долговременные изменения локального обилия видов птиц.
4. Провести анализ воздействия абиотических факторов (теплообеспеченности и количества осадков) на межгодовые флуктуации обилия фоновых видов птиц, а также оценить направленность и степень их влияния на долговременные изменения видовой плотности в горных условиях.
5. Оценить влияние фенологических сроков весеннего развития природы на формирование населения гнездящихся птиц.
6. Выявить влияние сроков прилета птиц и их долговременных изменений на формирование гнездового населения в горных условиях.аа
7. Установить экологические закономерности формирования и многолетних изменений локального гнездового и зимнего населения птиц в горных условиях.
Основные положения, выносимые на защиту.
1. Население птиц конкретного участка в горных условиях юга Восточной Сибири ежегодно формируется преимущественно из новых особей - иммигрантов. Этот процесс зависит от экологической обстановки, складывающейся в связи с условиями погоды весной и в начале лета, и соотношения процессов эмиграции и иммиграции. К главным факторам, обуславливающим формирование локального обилия вида, относятся фенологические и гидротермические особенности весны и начала лета текущего года.
2. В горных условиях Сибири в последние десятилетия среди дальних мигрантов преобладают виды с отрицательным трендом долговременных изменений гнездового обилия, а у оседлых видов и ближних мигрантов соотношение количества видов с положительными и отрицательными трендами примерно равное.
3. Сообщество птиц в горных условиях формируется и функционирует не по конкурентному, а по линдивидуалистическому типу. Изменения плотности разных видов (межгодовые флуктуации) обычно происходят независимо друг от друга. Асинхронный характер колебаний обилия у разных видов способствует стабилизации плотности населения в целом.
Научная новизна работы. Впервые проведены долгосрочные 25-летние стационарные исследования изменения гнездового и зимнего населения птиц в условиях гор юга Восточной Сибири, охватывающие лесной, подгольцово-субальпийский и гольцово-альпийский высотные пояса, которые позволили раскрыть закономерности процессов формирования локального обилия видов и определяющие их факторы. Рассмотрен в сравнительном плане современный состав авифауны горных систем юга Восточной Сибири, позволивший утверждать об орнитофаунистическом единстве этого региона, несмотря на различия в процессах фауногенезиса в его отдельных частях.
Раскрыты механизмы ежегодного формирования локального населения птиц под влиянием изменяющихся параметров местообитаний, в том числе под воздействием абиотических факторов теплообеспеченности и осадков, а также географического положения мест зимовок и складывающихся в них условий, позволившие по-новому определить специфику формирования населения птиц в горных условиях.
Выявлены определяющие факторы, трансформирующие структуру населения в горных условиях. В горно-лесных и высокогорных местообитаниях, при относительной стабильности структуры растительности и иных элементов местности, ведущее значение в процессах формирования локальной плотности видов птиц приобретают метеорологические характеристики сезона, оказывающие влияние на изменения доступных кормов и гнездопригодности территории. К главным факторам, обуславливающим формирование локального обилия вида, относятся фенологические и гидротермические особенности весны и начала лета текущего года.
Анализ долговременных изменений сроков прилета птиц, проведенный впервые в горных условиях Восточной Сибири, позволил выявить основные тренды их изменений, которые влияют на процессы формирования локального обилия видов.
Теоретическое и практическое значение. Выявленные закономерности формирования локального населения птиц в горных условиях позволяют расширить представление о механизмах популяционной динамики и формирования сообществ животных. Результаты исследований являются точкой отсчета для дальнейшего изучения динамических процессов в популяциях птиц и роли естественных воздействий на изменения сообществ птиц в будущем.
Собранные материалы, уникальные по объему и длительности наблюдений, имеют большое значение для работ по оценке различного рода воздействий на птиц в условиях горных систем юга Восточной Сибири, для оценки ответов биоты на глобальные изменения природной среды и климата, а также для организации мониторинга биоты на Участке всемирного наследия Озеро Байкал. Они послужили основой для организации и осуществления программы долговременного мониторинга в Баргузинском государственном природном биосферном заповеднике, в том числе в рамках программы ЮНЕСКО Человек и биосфера. Данные о распространении и численности редких и исчезающих видов птиц включены в последнее издание Красной книги Республики Бурятия (2005). Методические подходы, разработанные в диссертации, использованы при подготовке рекомендаций по организации комплексного экологического мониторинга состояния биоты в заповедниках и национальных парках Байкальской природной территории.
Полученные результаты используются в преподавании различных курсов на базе кафедры сервиса, туризма и рекреации Восточно-Сибирской государственной академии культуры и искусств (Биоэкология, Орнитология, Охрана природы, Заповедное дело, ООПТ Сибири, Биоразнообразие Байкальского региона), при проведении учебных и производственных практик студентов. Кроме того, автором было подготовлено и опубликовано учебное пособие Заповедное дело (2009) с грифом УМО учебных заведений РФ по образованию в области сервиса и туризма Минобрнауки России.
ичный вклад соискателя. Представленная диссертационная работа является обобщением результатов 25-летней работы автора. Автору принадлежит разработка программы исследований, выполнение полевых учетных работ во все сезоны года и фенологических наблюдений за прилетом птиц в 1983-2007 гг., камеральная обработка и обобщение материалов, интерпретация и публикация полученных результатов, формулировка и обоснование концепции, а также выводы.
Апробация работы. Материалы диссертации были доложены и обсуждены на более чем 40 международных, всесоюзных, российских и региональных конференциях и симпозиумах, в том числе: XI (Казань, 2001), XII (Ставрополь, 2006) и XIII (Оренбург, 2010) междунар. орнитол. конф. Северной Евразии; I, II, III и IV междунар. орнитол. конф. Современные проблемы орнитологии Сибири и Центральной Азии (Улан-Удэ, 2000, 2003, 2006, 2009); Междунар. симпоз. Многолетняя динамика численности птиц и млекопитающих в связи с глобальными изменениями климата (Казань, 2002); III конф. орнитологов Сибири (Томск, 1987); XII Прибалт. орнитол. конф. (Паланга, 1988); Всесоюз. совещ. по проблеме кадастра и учета животного мира (Уфа, 1989); Всесоюз. конф. Заповедники СССР - их настоящее и будущее (Новгород, 1990); регион. науч. конф. Биологическое разнообразие животных Сибири (Томск, 1998); междунар. науч. конф. Экология и рациональное природопользование на рубеже веков (Томск, 2000); совещ. Ключевые орнитологические территории России (Шушенское, 2000); регион. науч.-практич. конф. Структура и функционирование экосистем Байкальской Сибири (Улан-Удэ, 2002); Сибирская зоолог. конф. (Новосибирск, 2004); Всерос. конф. Природная и антропогенная динамика наземных экосистем (Иркутск, 2005); Междунар. конф. Проблемы популяционной экологии животных (Томск, 2006); II Междунар. науч. конф. Разнообразие почв и биоты Северной и Центральной Азии (Улан-Удэ, 2011).
Публикации. Автором опубликовано 242 печатных работы, из них по теме диссертации - 91, в том числе 3 монографии и 13 статей в рецензируемых изданиях, а также 1 учебное пособие.
Структура и объем диссертации. Работа состоит из введения, 7 глав, выводов, списка литературы, включающего 645 наименований, из них 126 работ на иностранных языках, содержит 86 таблиц и 30 рисунков. Общий объем диссертации 378 страниц.
Благодарности. Автор выражает глубокую признательность всем сотрудникам Баргузинского заповедника, принимавшим участие в проведении круглогодичных полевых работ на протяжении четверти века. Особой благодарности заслуживают несколько поколений сотрудников Баргузинского заповедника, которые на протяжении более 60 лет регистрировали даты весеннего прилета птиц. Без их самоотверженного труда такого рода обобщения были бы просто невозможны. Искренне благодарен своему учителю и наставнику С.С. Москвитину, всем коллегам, с которыми довелось вместе работать и общаться, сделавшим ряд ценных замечаний по отдельным публикациям, положенным в основу настоящей работы. Особую благодарность выражаю научному консультанту доктору биологических наук, профессору Ц.З. Доржиеву за ценные советы и критические замечания при подготовке рукописи диссертации.
Глава 1. Условия обитания птиц в горных системах юга Восточной Сибири
Приведена физико-географическая характеристика горных систем юга Восточной Сибири, включающих хребты с высокогорьями (с высотами свыше 2000 м над ур. м.) (Баргузинский, Хамар-Дабан, восточная часть Восточного Саяна с Тункинскими, Китойскими и Бельскими гольцами, Мунку-Сардыком, Большим Саяном, Окинским хребтом и Окинским плоскогорьем) и межгорные котловины (рис. 1).
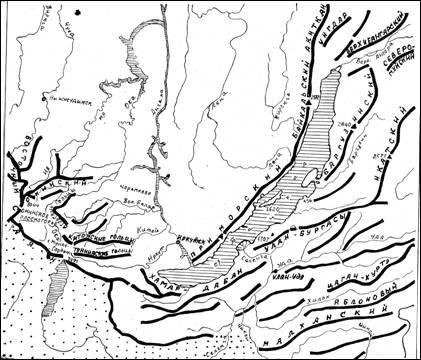
Рис. 1. Схема горных систем юга Восточной Сибири
Характерным отличием гор региона являются горные леса, смыкающиеся с таежными массивами предгорий Восточной Сибири (Бёме, Банин, 2001). На южных и восточных склонах с минимальными осадками развиты горные степи. Субальпийский пояс представлен верхней границей хвойных лесов с зарослями кедрового стланика и кустарниковых форм берез, ив и рододендронов. Субальпийские и альпийские луга занимают сравнительно небольшие площади, обычно на наиболее прогреваемых южных склонах. Большая часть плосковершинных участков хребтов и нагорных плато покрыта горными тундрами - кустарниковой, мохово-лишайниковой, каменистой и лишайниковой.
Более подробно представлено описание условий существования птиц в различных высотных поясах на ключевом участке исследований - на Баргузинском хребте и дана характеристика долговременных изменений климатических параметров на основе материалов наблюдений метеостанции Давша (Баргузинский хребет, побережье оз. Байкал).
Глава 2. Материал и методы исследований
Основой для данной работы послужили результаты изучения птиц гор юга Восточной Сибири несколькими поколениями орнитологов, а также собственные долговременные (1983-2007 гг.) исследования автора на Баргузинском Хребте, а также кратковременные обследования на Хамар-Дабане и Восточном Саяне.
Ключевой участок для выполнения программы долговременных орнитологических исследований находится на территории Баргузинского государственного природного биосферного заповедника (5401' - 5456' с.ш., 10928' - 11022' в.д.). Основополагающую часть использованных материалов составили 25-летние данные автора, полученные на строго фиксированных постоянных, предварительно промеренных и размеченных маршрутах и стационарных участках наблюдений, и дополненные исследованиями в 2008-2011 гг.
Летние учеты птиц выполнялись в период с 10 июня по 15 июля (первая половина лета) и с 16 июля по 20 августа (вторая половина лета) при двукратном проходе на строго фиксированных маршрутах. Зимние учеты птиц проводились ежегодно в период зимней стабилизации населения (субсезон морозной зимы, 15.01-28.02) и не ежегодно - в ноябре и декабре (субсезон снежной зимы).
Маршрутные учеты выполнялись без ограничения ширины трансекта, с последующим пересчетом на площадь по дальностям обнаружения интервальным методом (Равкин, 1967). Обоснование этой методики, техника проведения учетов и обработка данных в последнее время вновь детально изложены в работах Ю.С. Равкина с соавторами. (1999, 2007, 2008, 2009). Видовые названия птиц приводятся по Л.С. Степаняну (2003). Результаты учетов, выполненных в первую половину лета (10 июня - 15 июля), условно соответствуют гнездовому населению птиц. Дополнительные учеты выполнялись во все сезоны года во время экспедиционных обследований различных участков Баргузинского хребта, от побережья Байкала до гольцово-альпийского пояса.
Динамика численности летнего населения птиц прослежена в 1984-2007 гг. на постоянных учетных маршрутах, расположенных в долинах трех рек от побережья оз. Байкал до высокогорий Баргузинского хребта (460-1700 м над у. м.). На этих трансектах ключевого участка общей протяженностью 104,8 км, разбитых на 11 выделов, представлена большая часть разнообразия местообитаний нижней и верхней части горно-лесного и подгольцово-субальпийского поясов (Ананин, 2006, 2010). В данной работе мы отказались от биотопического разделения населения птиц и рассматривали группировки птиц на конкретных пространственных участках.
Постоянный маршрут по долине р. Езовка (КУ-Е), общей протяженностью 35,4 км, включает три участка:
ПР-Е - прибрежно-равнинный (10,6 км, 460-470 м над у. м.),
ПГ-Е - предгорный (13,2 км, 470-615 м над у. м.),
ГЛ-Е - горно-лесной (11,6 км, 615-1150 м над у. м.).
Постоянный маршрут по долине р. Большая (КУ-Б), общей протяженностью 41,9 км, разделен на четыре фрагмента:
ПР-Б - прибрежно-равнинный (10,8 км, 460-520 м над у. м.),
Р-Б - равнинный (8,9 км, 520-560 м над у. м.),
РМ-Б - равнинно-моренный (10,7 км, 560-580 м над у. м.),
ПГ-Б - предгорный (11,5 км, 580-630 м над у. м.).
Постоянный маршрут по долине р. Давша (КУ-Д), длиной 27,5 км, разбит на четыре участка:
ПР-Д - прибрежно-равнинный (10,5 км, 470-515 м над у. м.),
ПГ-Д - предгорный (6,3 км, 515-720 м над у. м.),
ГЛ-Д - горно-лесной (6,7 км, 720-1280 м над у. м.),
ПС-Д - подгольцово-субальпийский (4,0 км, 1280-1700 м над у. м.).
Общая протяженность пеших маршрутных учетов - 15200 км, в том числе летом - 6100 км, зимой - 7300 км, из них на постоянных маршрутах соответственно 4474 и 1556 км (1984-2007 гг.).
Основную часть использованных материалов по фенологии прилета птиц составили 25-летние данные автора, полученные в 1983-2007 гг. на постоянных маршрутах и стационарных участках наблюдений в зоне прибайкальских террас, на расстоянии до 10Ц15 км от озера Байкал. Для анализа сроков весеннего прилета птиц по первому появлению не включались данные по видам, отмеченным в год наблюдений зимой. Собраны сведения о весенних миграциях 124 видов птиц.
Помимо собственных материалов в работе использованы наблюдения научных сотрудников и работников охраны заповедника, относящиеся, прежде всего, к единичным регистрациям массовых и редких индикаторных видов. Дополнительно привлечены архивные материалы по фенологии весеннего прилета птиц и Летописи природы Баргузинского государственного заповедника за период с 1938 по 1983 г.
Статистические расчеты реализованы с применением пакетов программ Statistica 6.0 и Excel с учетом методических рекомендаций, предложенных для анализа временных рядов данных (Коросов, 2007). Кроме специально оговоренных случаев, использованы методы непараметрической статистики. Для оценки силы связи применялся ранговый коэффициент корреляции Кендалла (r?) или Спирмена (rS) (Песенко, 1982). В качестве меры общности двух объектов использован коэффициент Жаккара (Jaccard, 1902). Для оценки сходства (различия) облика населения птиц использован коэффициент Жаккара в модификации Наумова для количественных признаков (Наумов, 1964). Дополнительно, в специально оговоренных случаях, для качественного сравнения видового состава орнитокомплексов использован индекс Чекановского-Съеренсена (Песенко, 1982).
Долговременная тенденция изменения обилия птиц на участках определялась по коэффициенту детерминации в регрессионном уравнении, а его достоверность - по F-критерию (Коросов, 2007).
Глава 3. Орнитологическая изученность и современный состав орнитофауны горных систем юга Восточной Сибири
3.1. История орнитофаунистических исследований горных систем юга Восточной Сибири.
Исследования фауны птиц горных систем юга Восточной Сибири охватывают период около 250 лет, в результате чего современная авифауна оказалась относительно хорошо изученной. Историю ее изучения можно проследить в нескольких специальных работах (Гагина, 1960а; Измайлов, Хабаева, Боровицкая, 1973; Измайлов, 1967; Измайлов, Боровицкая, 1973, 1982; Скрябин, 1975; Ефимов и др., 1982; Толчин, 1982; Доржиев, 1997, 2003; Ананин, 2000; Овдин, Разуваев, 2003; Доржиев, Малеев, 2011).
Первый этап завершился к концу XIX в. обобщающим двухтомным трудом В.К. Тачановского о птицах Восточной Сибири (Taczanowski, 1891-1893). Следующий период, до середины ХХ столетия, характеризуется углубленными стационарными исследованиями сотрудников Баргузинского заповедника на Баргузинском хребте (Скрябин, Филонов, 1962), и экспедиционными работами в других частях региона с опубликованием списка птиц Восточной Сибири (Козлова, 1930; Скалон, 1936; Гагина, 1961).
Третий период (1960-1980-ые гг.) ознаменован продолжением изучения птиц Баргузинского хребта с участием сотрудников Баргузинского заповедника (Беляев К.Г., Жаров В.Р., Федоров А.В.), специалисты вузов региона и СО АН СССР (Ануфриев В.М., Баранов А.А., Богородский Ю.В., Боровицкая Г.К., Доржиев Ц.З., Измайлов И.В, Кельберг Г.В., Ким Т.А., Морошенко Н.В., Подковыров В.А., Пыжъянов С.В., Садков В.С., Сафронов Н.Н., Юмов Б.О.) Стационарные орнитофаунистические исследования на хребте Хамар-Дабан связаны с организацией там Байкальского государственного заповедника и завершились опубликованием сводки Птицы Хамар-Дабана (Васильченко, 1987).
Современный этап авифаунистических изысканий характеризуется широким применением методов количественного учета населения и детальными исследованиями экологии отдельных видов птиц орнитологами Бурятии под руководством Ц.З. Доржиева (Ананин А.А., Дашинимаев В.М, Елаев Э.Н., Ешеев В.Е., Мункуева Н.А., Разуваев А.Е.) и г. Иркутска (Дурнев Ю.А., Попов В.В., Рябцев В.В., Сонина М.В.).
3.2. Орнитофаунистическая характеристика горных систем юга Восточной Сибири
Видовой состав птиц горных систем юга Восточной Сибири исследован с использованием доступных нам орнитологических публикаций. Современная орнитофауна региона достаточно разнообразна, составляет 47,9% орнитофауны России (Коблик и др., 2006) и 93,3 % фауны птиц территории российской части бассейна озера Байкал (Доржиев, 2011). В горных системах юга Восточной Сибири зарегистрировано 378 видов птиц, из них 253 вида (66,9 %) - гнездящиеся (в том числе 54 (14,3 %) - оседлые), пролетных видов - 44 (11,7 %) и залетных - 78 видов (20,6 %) (табл. 1).
Таблица 1
Распределение фауны птиц горных систем юга Восточной Сибири по характеру пребывания
Характер пребывания |
Баргузинский хребет |
Хамар-Дабан |
Восточный Саян |
Горные системы юга Восточной Сибири |
||||
абс. |
% |
абс. |
% |
абс. |
% |
абс. |
% |
|
Всего видов: |
321 |
100,0 |
285 |
100,0 |
330 |
100,0 |
378 |
100,0 |
Гнездящиеся и вероятно гнездящиеся |
215 |
67,0 |
169 |
59,3 |
218 |
66,1 |
253 |
66,9 |
аа Из них оседлые |
46 |
14,3 |
60 |
21,1 |
56 |
17,0 |
54 |
14,3 |
Пролетные |
45 |
14,0 |
96 |
33,7 |
48 |
14,6 |
44 |
11,7 |
етующие |
- |
- |
1 |
0,3 |
8 |
2,4 |
- |
- |
Зимующие |
3 |
0,9 |
2 |
0,7 |
7 |
2,1 |
3 |
0,8 |
Залетные |
58 |
18,1 |
17 |
6,0 |
49 |
14,8 |
78 |
20,6 |
В систематическом отношении орнитофауна исследуемого района довольно разнообразна, но доля участия в ней разных таксономических групп весьма неравнозначна. Наибольшим числом видов представлен отряд Passeriformes (173 вида, 45,8 % от общего числа видов, из них гнездящихся 134 вида). Затем следуют такие крупные отряды как Charadriiformes (67 видов, 17,7 %), Anseriformes (33 вида, 8,7 %) и Falconiformes (32 вида, 8,5 %). Все остальные отряды насчитывают от 1 до 12 видов (табл. 2).
Таблица 2
Таксономическая структура авифауны горных систем юга Восточной Сибири
Отряд |
Баргузинский хребет |
Хамар-Дабан |
Восточный Саян |
Горные системы юга Восточной Сибири |
||||
Nобщ |
Nгнез |
Nобщ |
Nгнез |
Nобщ |
Nгнез |
Nобщ |
Nгнез |
|
Gaviiformes |
3 |
2 |
2 |
0 |
2 |
1 |
3 |
2 |
Podicipediformes |
4 |
4 |
4 |
0 |
5 |
3 |
5 |
4 |
Pelecaniformes |
1 |
1 |
1 |
0 |
1 |
0 |
1 |
1 |
Ciconiiformes |
4 |
3 |
4 |
1 |
4 |
3 |
5 |
3 |
Phoenicopteriformes |
1 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
Anseriformes |
31 |
20 |
26 |
7 |
29 |
14 |
33 |
20 |
Falconiformes |
28 |
18 |
26 |
12 |
30 |
17 |
32 |
21 |
Galliformes |
9 |
9 |
9 |
9 |
12 |
10 |
12 |
10 |
Gruiformes |
10 |
4 |
5 |
0 |
10 |
5 |
12 |
5 |
Charadriiformes |
61 |
24 |
45 |
12 |
44 |
15 |
67 |
25 |
Columbiformes |
4 |
4 |
3 |
3 |
6 |
4 |
6 |
4 |
Cuculiformes |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
Strigiformes |
10 |
8 |
10 |
9 |
12 |
9 |
12 |
9 |
Caprimulgiformes |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
Apodiformes |
3 |
3 |
3 |
3 |
3 |
3 |
3 |
3 |
Coraciiformes |
1 |
1 |
1 |
1 |
1 |
0 |
1 |
1 |
Upupiformes |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
Piciformes |
7 |
7 |
7 |
7 |
7 |
7 |
7 |
7 |
Passeriformes |
139 |
103 |
134 |
100 |
159 |
123 |
173 |
134 |
ВСЕГО: |
321 |
215 |
285 |
169 |
330 |
218 |
378 |
253 |
Nобщ - общее число видов, Nгнез - число гнездящихся видов.
При выделении авифаунистических комплексов за основу было взято орнитогеографическое деление Палеарктики Б.К. Штегмана (1938) с дополнением (Дементьев, 1937; Гагина, 1960) и некоторыми уточнениями Ц.З. Доржиева (Доржиев, 1997), А.А. Баранова (2007) и В.Г. Бабенко (Бабенко, Фадеева, 2010).
Структура гнездящейся орнитофауны горных систем юга Восточной Сибири включает 9 фаунистических элементов. Если не принимать во внимание виды, центры происхождения которых к настоящему времени недостаточно ясны (группы голарктических, транспалеарктических и других широкораспространенных видов, общее число которых составляет 79, а доля - 31,2 %), можно указать, что основу авифауны региона составляют виды трех комплексов - сибирского (25,3 %), европейского (14,2 %) и китайского (11,5 %). Довольно существенен вклад монгольского (8,3 %) и тибетского (5,5 %) комплексов. Значение других типов фауны - арктического (3,2 %) и средиземноморского (0,8 %) в формировании орнитофауны исследуемого района невелико (табл. 3).
Таблица 3
Распределение гнездящихся видов птиц горных систем юга Восточной Сибири по типам фауны
Тип фауны |
Баргузинский хребет |
Хамар-Дабан |
Восточный Саян |
Горные системы юга Восточной Сибири |
||||
абс. |
% |
абс. |
% |
абс. |
% |
абс. |
% |
|
Арктический |
6 |
2,8 |
4 |
2,4 |
2 |
0,9 |
8 |
3,2 |
Сибирский |
61 |
28,3 |
52 |
30,7 |
53 |
24,3 |
64 |
25,3 |
Монгольский |
14 |
6,5 |
10 |
5,9 |
18 |
8,3 |
21 |
8,3 |
Китайский |
26 |
12,1 |
25 |
14,8 |
27 |
12,4 |
29 |
11,5 |
Тибетский |
9 |
4,2 |
9 |
5,3 |
13 |
6,0 |
14 |
5,5 |
Европейский |
26 |
12,1 |
21 |
12,4 |
33 |
15,1 |
36 |
14,2 |
Средиземноморский |
1 |
0,5 |
2 |
1,2 |
2 |
0,9 |
2 |
0,8 |
Транспалеаркты |
58 |
27,0 |
41 |
24,3 |
56 |
25,7 |
60 |
23,7 |
Голарктический |
5 |
2,3 |
2 |
1,2 |
4 |
1,8 |
5 |
2,0 |
Широко-распространенные |
9 |
4,2 |
3 |
1,8 |
10 |
4,6 |
14 |
5,5 |
ВСЕГО: |
215 |
100,0 |
169 |
100,0 |
218 |
100,0 |
253 |
100,0 |
В структуре фауны гнездящихся видов птиц Баргузинского хребта, по сравнению с общей гнездовой орнитофауной горных систем юга Восточной Сибири, более высокую долю составляют представители сибирского (28,3 %) и китайского (12,1 %) типов фауны. На хребте Хамар-Дабан отмечается сходная картина - доля видов сибирского и китайского типа фаун составляет, соответственно, 30,5 и 15,0 %. В восточной части Восточного Саяна, по сравнению с орнитофаунами хребтов Хамар-Дабан и Баргузинский, отмечено превышение доли европейского (15,2 %), монгольского (8,3 %) и тибетского (6,0 %) типов фауны. Эти отличия определяются географическим положением сравниваемых участков и особенностями их природных условий.
По характеру современного распространения гнездящихся видов в орнитофауне исследуемого района подразделяется несколько орнитофаунистических комплексов. Выделение таких комплексов выполнено на основе подходов, сформулированных Ц.З. Доржиевым (Доржиев, Елаев, 2000) с изменениями и дополнениями на основе работ В.А. Нечаева (Нечаев, 1991), В.С. Жукова (Жуков, 2004) и В.Г. Бабенко (Бабенко, Фадеева, 2010). В фауне птиц горных систем юга Восточной Сибири преобладают палеарктические виды (105 видов, 41,5 %). На долю голарктических видов приходится 17,0 % (43 вида). Виды, имеющие всесветное распространение (космополиты), составляют 9,5 % (24 вида). Азиатский комплекс представлен 79 видами (31,3 %). Среди них наибольшее значение имеют североазиатская (30 видов, 11,9 %) и центральноазиатская (21, 8,3 %) подгруппы, менее всего представлены восточноазиатская (16, 6,3 %) и трансазиатская (12 видов, 4,8 %) подгруппы (табл. 4).
Таблица 4
Распределение гнездящихся видов птиц горных систем юга Восточной Сибири по типам ареалов
Орнитофаунистические комплексы |
Баргузинский хребет |
Хамар-Дабан |
Восточный Саян |
Горные системы юга Восточной Сибири |
||||
абс. |
% |
абс. |
% |
абс. |
% |
абс. |
% |
|
Космополиты |
24 |
11,2 |
14 |
8,3 |
20 |
9,2 |
24 |
9,5 |
Голарктическая группа |
41 |
19,1 |
27 |
16,0 |
36 |
16,5 |
43 |
17,0 |
Палеарктическая группа |
88 |
40,9 |
67 |
39,6 |
92 |
42,2 |
105 |
41,5 |
Азиатско-американская группа |
2 |
0,9 |
1 |
0,6 |
0 |
0 |
2 |
0,8 |
Азиатская группа |
60 |
27,9 |
60 |
35,5 |
70 |
32,1 |
79 |
31,2 |
Трансазиатская подгруппа |
12 |
5,6 |
12 |
7,1 |
12 |
5,5 |
12 |
4,7 |
Североазиатская подгруппа |
28 |
13,0 |
23 |
13,6 |
22 |
10,1 |
30 |
11,9 |
Центральноазиатская подгруппа |
5 |
2,3 |
11 |
6,5 |
21 |
9,6 |
21 |
8,3 |
Восточноазиатская подгруппа |
15 |
7,0 |
14 |
8,3 |
15 |
6,9 |
16 |
6,3 |
ВСЕГО: |
215 |
100,0 |
169 |
100,0 |
218 |
100,0 |
253 |
100,0 |
Сопоставление орнитофаун Баргузинского хребта, хребта Хамар-Дабан и восточной части Восточного Саяна, основных высокогорных подразделений горных систем юга Восточной Сибири, выполнено на основе коэффициента фаунистического сходства, рассчитанного по формуле Чекановского-Съеренсена с дополнениями и изменениями Н.И. Лариной и Г.М. Гурылевой (1972).
Сравнение фаунистического сходства полных локальных орнитофаун показывает очень близкие значения коэффициента сходства между всеми тремя участками (85,7 - 87,1 %). При сравнении фаун гнездящихся видов максимальное сходство зарегистрировано между наиболее удаленными друг от друга Баргузинским хребтом и Восточным Саяном (84,1 %, 182 общих вида), а минимальное - между орнитофаунами Хамар-Дабана и Баргузинского хребта (79,2 %, 152 общих вида) (табл. 5). Такой высокий уровень фаунистического сходства недостаточен для обособления сообществ в ранге самостоятельных зоогеографических единиц (Ларина, Гурылева, 1972) и подтверждает орнитофаунистическое единство горных систем юга Восточной Сибири.
Таблица 5
Сравнение локальных орнитофаун горных систем юга Восточной Сибири
Сравниваемые орнитофауны |
Коэффициент фаунистического сходства орнитофаун (%) / число видов птиц, общих для двух сравниваемых участков |
|
полные |
гнездовые |
|
Баргузинский хребет - Хамар-Дабан |
87,1 / 264 |
79,2 / 152 |
Баргузинский хребет - Восточный Саян |
85,7 / 279 |
84,1 / 182 |
Хамар-Дабан - Восточный Саян |
86,3 / 265 |
80,1 / 155 |
Таким образом, орнитофауна горных систем юга Восточной Сибири достаточно разнообразна (378 видов, в том числе 253 гнездящихся), ее основу составляют, помимо широкораспространенных видов, представители сибирского, европейского, китайского и монгольского типов фаун, а по ареалогической структуре - виды палеарктической, голарктической и азиатской групп. Среди последних максимально представлены североазиатская и центральноазиатская подгруппы, а в меньшей степени - виды восточноазиатской и трансазиатской подгрупп. Авифаунистическое сходство между отдельными горными системами очень велико, подтверждая единство фауны птиц региона, несмотря на различия в процессах фауногенезиса в его отдельных частях.
Глава 4. Пространственная структура Населения птиц горных систем юга Восточной Сибири
(на примере Баргузинского хребта)
4.1. Пространственное распределение птиц в летний период
На ключевом участке исследований по составу населения и обилию особей можно обособить орнитокомплексы прибрежно-равнинного, предгорного, горно-лесного, подгольцово-субальпийского и гольцово-альпийского выделов. Каждый из ландшафтных поясов характеризуется специфичным обликом населения птиц, отражающим основные особенности их природных условий.
Для ландшафтов как Баргузинского хребта, так и всех остальных горных систем юга Восточной Сибири характерна низкая численность птиц (Кельберг, 1985; Ананин, 2006), что объясняется в первую очередь невысокой суммарной продуктивностью фитоценозов и относительно низким количеством всех доступных субстратов (Юдкин, 2002, 2009), обусловленным особенностями растительного покрова региона.
С повышением высоты местности видовое богатство постепенно убывает от 103 на прибайкальских террасах (прибрежно-равнинные выделы) до 44 в гольцово-альпийском поясе, всего на стационаре в гнездовой период зарегистрированы 132 вида. Самая высокая среднемноголетняя (усредненная за 1984-2007 гг.) плотность населения птиц в первую половину лета наблюдалась в предгорьях (нижняя часть горно-лесного пояса) (273,5 особ./км2), где климатические условия для западного макросклона Баргузинского хребта наиболее оптимальны.
Второй максимум плотности населения (268,9 особ./км2) зафиксирован в подгольцово-субальпийском поясе. В целом, с повышением высоты местности обилие птиц сокращается (рис. 2).

Рис. 2. Распределение среднемноголетней плотности населения птиц высотных поясов западного макросклона Баргузинского хребта в I и II половину лета (1984Ц2007 гг. особ./км2 ): ПР - прибрежно-равнинный пояс; ПГ - предгорный пояс (нижняя часть горно-лесного пояса); ГЛ - верхняя часть горно-лесного пояса; ПС - подгольцово-субальпийский пояс; ГА - гольцово-альпийский пояс.
Во вторую половину лета распределение населения птиц по высотному профилю имеет сходный характер и существенно не изменяется. По сравнению с гнездовым периодом во вторую половину лета обилие птиц возрастает в предгорьях (на 16,8%) и в верхней части горно-лесного пояса (на 11,8%). В остальных высотных выделах во вторую половину лета наблюдались откочевки птиц после завершения гнездования.
Анализ структуры гнездового населения птиц ключевого участка демонстрирует закономерное распределение доли разных типов фаун в зависимости от высоты местности и преобладающего типа растительности. Участие сибирских (таежных) видов максимально в темнохвойных лесах - в горно-лесном поясе (61,3%), европейских - в предгорьях (6,7%), где заметна доля мелколиственных фрагментов лесов, а тибетских, монгольских и арктических видов - в гольцово-альпийском поясе (соответственно, 10,3, 7,7 и 5,1%), транспалеарктов - в прибрежно-равнинном выделе (36,1%) с высокой долей водоемов и пойменных местообитаний. Китайские по происхождению виды птиц, тяготеющие к лугам, болотам и приручьевым смешанным лесам, распределены примерно одинаково во всех высотных поясах.
В результате 25-летних исследований на ключевом участке выявлены орнитокомплексы прибрежно-равнинного, предгорного, горно-лесного, подгольцово-субальпийского и гольцово-альпийского выделов, достаточно обособленные по составу населения и обилию особей. На Баргузинском хребте отмечается постепенное снижение видового разнообразия и суммарного обилия видов птиц к верхним высотным поясам, аналогичное наблюдениям на Хамар-Дабане (Кельберг, 1985) и в других южных горных системах (Матюшкин, 1966; Равкин, 1973; Цыбулин, 1999), что коррелирует с упрощением ярусной структуры растительных сообществ по мере увеличения высоты местности.
4.2. Пространственное распределение птиц в зимний период
В зимние периоды 1984/85-2006/07 гг. на ключевом участке отмечен 31 вид птиц 5 отрядов. Доминирует буроголовая гаичка, к субдоминантам относятся московка и обыкновенный поползень, а в горно-лесном выделе к ним добавляется обыкновенная чечетка. 7 видов многочисленны, 7 - обычны, остальные малочисленны и редки. Максимальное среднемноголетнее обилие зимующих птиц - в горно-лесном поясе (341,0 особ./км2). Оно снижается в предгорных и равнинных лесах до 176 особ./км2 (рис. 3).
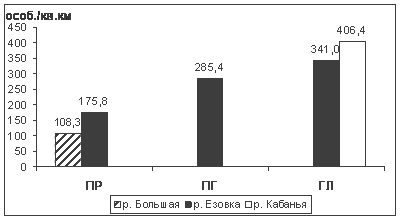
Рис. 3. Распределение среднемноголетней плотности (особ./км2 ) зимнего населения птиц высотных поясов ключевого участка Баргузинского хребта (1984/1985 - 2006/2007 гг.): ПР - прибрежно-равнинный пояс; ПГ - предгорный пояс (нижняя часть горно-лесного пояса); ГЛ - верхняя часть горно-лесного пояса.
На прибрежно-равнинном выделе зарегистрировано 22 вида. На этом участке доминируют буроголовая гаичка и поползень. Кроме них к многочисленным видам относятся длиннохвостая синица, московка, пестрый дятел и белокрылый клест. Обычны обыкновенная чечетка, кедровка, рябчик, кукша, щур, пищуха и трехпалый дятел. Остальные виды малочисленны и редки.
В нижней части горно-лесного пояса (предгорный выдел) отмечены 23 вида. В этом орнитокомплексе абсолютно доминирует буроголовая гаичка. Субдоминанты - обыкновенный поползень и московка. Многочисленны пестрый дятел, длиннохвостая синица, кедровка и белокрылый клест. В состав обычных видов нижней части горно-лесного пояса дополнительно включается желтоголовый королек.
В верхней части горно-лесного пояса, по сравнению с зимним населением птиц предгорного выдела, к числу субдоминантов присоединяется обыкновенная чечетка. Многочисленны 5 видов. К числу обычных видов (11) в этом выделе добавляются желна, обыкновенный клест и не ежегодно зимующий серый снегирь.
Следовательно, минимальное обилие и разнообразие зимующих птиц регистрируется в прибрежно-равнинном выделе, оно повышается по мере продвижения к верхней части горно-лесного и подгольцово-субальпийскому поясу. К лидирующим по обилию видам в зимних орнитокомплексах относятся буроголовая гаичка, московка, обыкновенный поползень и обыкновенная чечетка.
Глава 5. Влияние экологических факторов на Многолетнюю динамику летнего обилия птиц в горных условиях
5.1. Динамика обилия фоновых видов птиц в летний период
Гнездовая плотность каждого вида птиц на ключевом участке Баргузинского хребта за 25-летний период исследований испытывала колебания различной амплитуды и длительности. Для большинства регулярно гнездящихся видов установлены кратковременные непрерывные флуктуации обилия с амплитудой, не превышающей 2Ц8-кратных значений.
Высокая изменчивость выявлена для белокрылого клеста, пеночки-зарнички, синехвостки и длиннохвостой синицы (13Ц19-кратные изменения), обыкновенной пищухи и пятнистого сверчка (27Ц37-кратные), а наибольшая - для вьюрка, склонного к значительным ежегодным перемещениям в пределах гнездового ареала (52-кратные).
Долговременные тенденции динамики плотности - стабильность, снижение или рост за период наблюдений - отражают изменения среды обитания на конкретном участке или общие тенденции динамики численности вида в ареале. Из 55 фоновых видов статистически достоверная тенденция увеличения обилия (положительный тренд) обнаружена у 8: чижа, рябчика, бурой пеночки, белокрылого клеста, желтоголового королька, певчего сверчка, пестрого дрозда и черныша (рис.4).
Статистически значимый отрицательный тренд зафиксирован для 16 видов (рис. 5): буроголовой гаички, пеночки-зарнички, горной трясогузки, обыкновенной чечевицы, овсянок: рыжей, желтобровой, седоголовой, белошапочной и дубровника, синего соловья, длиннохвостой синицы, сибирской чечевицы, сибирской мухоловки, краснозобого и оливкового дроздов, а также щура. Еще у 31 вида статистически существенные тренды не выявлены.
Раздельный анализ трендов изменения видового обилия на отдельных выделах показал, что статистически значимые тренды обнаруживаются не только у тех птиц, для которых они выявлены для всего участка. При этом для некоторых видов на разных выделах могут вскрываться разнонаправленные статистически существенные тренды.
|
|


Рис. 4. Многолетние изменения летней плотности видов птиц ключевого участка с положительным трендом (1984Ц2007 гг., I половина лета, особ./км2)
|
|
|
|
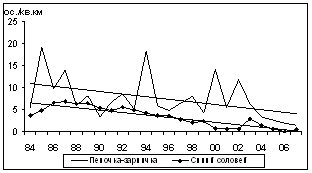
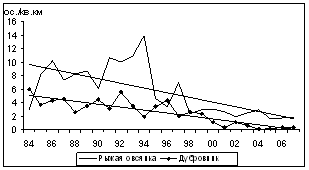

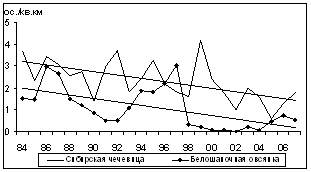
Рис. 5. Многолетние изменения летней плотности видов птиц ключевого участка с отрицательным трендом (1984Ц2007 гг., I половина лета, особ./км2)
Разнонаправленность трендов долговременного изменения обилия может свидетельствовать о наличии избирательного ежегодного заселения выделов под влиянием каких-то факторов, которое приводит к перераспределению видовой плотности между высотно-поясными участками или однотипными выделами в соседних речных долинах.
Оптимальные для вида местообитания характеризуются максимальной среднемноголетней плотностью и минимальной межгодовой изменчивостью видового обилия, которая оценивается коэффициентом вариации (CV, %). Выделы, на которых многолетняя динамика обилия вида тесно коррелирует друг с другом, но слабо связана с изменениями обилия на других участках, были объединены в корреляционные плеяды (Терентьев, 1959), либо обозначены как участки, на которых этот признак изменяется статистически независимо от других выделов. Большое количество выделенных корреляционных плеяд указывает на низкое сходство долговременных изменений обилия вида на постоянных участках наблюдений и на их относительно независимый характер.
По характеру ежегодного распределения по участкам 12 лидирующих по обилию видов разделились на две группы (табл. 6). Первая характеризуется достаточно синхронными изменениями видового обилия на отдельных участках наблюдения и формированием корреляционных плеяд с положительной связью. Она подразделяется на три подгруппы. В первую из них включены 6 видов (обыкновенный поползень, московка, буроголовая гаичка, корольковая пеночка, синехвостка и чиж), которые формируют 2Ц3 корреляционные плеяды с положительной корреляцией. Основная часть видового населения выделов объединена в единую плеяду, а 1Ц2 участка имеют население со статистически независимым характером межгодовых изменений плотности. Вторая подгруппа содержит только один вид (горная трясогузка), долговременная динамика обилия которого подразделяется на 6 корреляционных плеяд с относительно независимым характером межгодовых изменений.
Обособленное положение занимает пеночка-зарничка, для которой характерен статистически независимый характер долговременной динамики гнездовой плотности в одном из оптимальных местообитаний, а также в явно субоптимальных выделах, где вид регистрировался не ежегодно. Остальные участки, оптимальные для вида, с высокой и относительно стабильной плотностью гнездования, формируют отдельную корреляционную плеяду с многолетними синхронными изменениями обилия.
Вторая группа (пятнистый конек, зеленая пеночка, рябчик и пестрый дятел) характеризуется асинхронным типом динамики гнездовой плотности, который сопровождается межгодовым перераспределением видового обилия в пределах разных высотно-поясных выделов и между соседними речными долинами, что находит свое выражение в формировании корреляционных плеяд с отрицательной корреляцией. При этом следует учитывать, что многолетние изменения обилия в разных выделах у них происходят статистически независимо.
Таким образом, среди 55 исследованных видов за 25-летний период наблюдений у 8 выявлен положительный тренд долговременных изменений, у 16 - отрицательный и у 31 вида тренд статистически не значим. Избирательное заселение птицами разнокачественной среды приводит к существенным различиям в характере межгодовой изменчивости обилия в оптимальных и субоптимальных местообитаниях. По особенностям межгодовых флуктуаций виды подразделяются на две группы: с синхронным и асинхронным типом динамики плотности гнездования на высотно-поясных выделах. Для птиц с асинхронными изменениями локальной численности характерно перераспределение видового обилия между выделами одного высотного трансекта или между соседними речными долинами. а
Таблица 6
Группы участков со сходным характером долговременных изменений видового обилия,
выявленные методом корреляционных плеяд (1984Ц2007 гг.)
Вид |
Корреляционные плеяды |
||||||||||
с положительной корреляцией (p < 0,05) |
с отрицательной корреляцией (p < 0,05) |
||||||||||
Обыкн. поползень |
гл-д |
воу |
|
|
|
|
|
|
|
|
|
Московка |
пг-д, гл-д |
пр-е |
воу |
|
|
|
|
|
|
|
|
Буроголовая гаичка |
ра-б |
пг-б |
воу |
|
|
|
|
|
|
|
|
Корольковая пеночка |
пг-д, гл-д |
воу |
|
|
|
|
|
|
|
|
|
Синехвостка |
гл-е |
воу |
|
|
|
|
|
|
|
|
|
Пятнистый конек |
рм-б, гл-е |
пг-б, пг-е |
ра-б, |
пр-е |
пр-д |
пг-д |
гл-д |
пс-д |
|
пр-д, г-е, гл-д |
рм-б, пр-д |
Чиж |
пр-б |
гл-д |
воу |
|
|
|
|
|
|
|
|
Зеленая пеночка |
пр-б, гл-е |
пг-б, пр-е |
ра-б |
рм-б |
пг-е |
пр-д |
пг-д |
гл-д |
|
ра-б, пр-е, гл-е |
|
Пеночка-зарничка |
ра-б |
пс-д |
гл-д |
воу |
|
|
|
|
|
|
|
Рябчик |
пр-е, пг-е |
пр-б |
ра-б |
рм-б |
пг-б |
гл-е |
пр-д |
пг-д |
гл-д |
ра-б, гл-д |
рм-б, гл-е |
Горная трясогузка |
пр-б, гл-д |
ра-б, рм-б, пг-б, пр-е, пс-д |
пг-е |
гл-е |
пр-д |
пг-д |
|
|
|
|
|
Пестрый дятел |
пр-е, гл-е, пг-д |
ра-б |
гл-д |
воу |
|
|
|
|
|
пг-б, гл-д |
|
Примечания: Участки: ПР - прибрежно-равнинный, РА - равнинный, РМ - равнинно-моренный, ПГ - предгорный, ГЛ - горно-лесной, ПС - подгольцово-субальпийский; ЦБ - долина р. Большая, ЦЕ а - долина р. Езовка, ЦД - долина р. Давша; ВОУ - все остальные участки.
5.2. Влияние мест и условий зимовок на долговременные изменения локального обилия видов птиц
На ключевом участке в последнее десятилетие проявилась тенденция снижения плотности гнездования всего населения во всех исследованных выделах, что явилось следствием превышения доли видов с отрицательными трендами над числом видов с положительным трендом (рис. 6).
Отрицательные тренды долговременных изменений гнездовой плотности преобладают у дальних мигрантов (11 видов против 4 с положительными трендами). У оседлых видов, как и у ближних мигрантов, это соотношение равное (3:3 и 1:2, соответственно). Возросла доля видов с отрицательными трендами среди дальних мигрантов, зимующих на всех южно-азиатских зимовках: пакистано-индийской, китайской, индокитайской и в меньшей степени на филлипино-малакко-индонезийской, что может свидетельствовать об их неблагополучном состоянии. Такой же точки зрения в отношении причин снижения численности хищных птиц региона придерживается В.В. Рябцев (2009, 2010).

Рис. 6. Динамика плотности летнего населения птиц ключевого участка Баргузинского хребта (1984Ц2007 гг., I половина лета, особ./км2): пунктирная линия - среднемноголетний уровень обилия населения птиц на ключевом участке.
Детальные исследования (анализ ключевых факторов и выживаемости) на европейских птицах показали, что причинами самых резких снижений их численности были в ряде случаев не нарушения процесса размножения, а снижение выживаемости птиц на территориях зимовок, в особенности африканских (Enemar et al., 1984, 1994; Бурский, 1993, 2007; Morozov, 1993; Соколов, 1999 и др.). Эти заключения справедливы и в отношении южно-азиатских зимовок. В современный период для этих территорий характерно повышение уровня антропогенного пресса на местообитания вследствие огромной концентрации населения и возрастания степени загрязнения природной среды различными ядохимикатами, а также осуществление массового отлова мелких воробьиных птиц местным населением в период миграций и на зимовках с целью традиционного использования в пищу.
Выполненная нами классификация видов птиц по сходству долговременной динамики гнездовой плотности методом кластер-анализа, показала, что зимующие в пределах Баргузинского хребта гнездящиеся виды распределились, в основном, по трем группам, в одну из которых включены синицы, обитающие в стаях, часто совместных (московка, буроголовая гаичка, длиннохвостая синица), а также редко зимующий в этом районе чиж. Второй кластер образовали оседлые виды, чаще живущие в зимний период парами или небольшими семейными группами с общим достаточно высоким уровнем обилия (обыкновенный поползень, пестрый дятел, рябчик и белокрылый клест). Оседлые виды с более низким уровнем обилия (кедровка, кукша, обыкновенная пищуха и желтоголовый королек) составили третье объединение. Щур сформировал особый отдельный кластер, достаточно независимый от других групп птиц.
Дальние (тропические) мигранты составили 6 кластеров. В первый из них вошли краснозобый дрозд, сибирский жулан, седоголовая овсянка, соловей-красношейка, пятнистый сверчок, бурая и корольковая пеночки, имеющие очень широкую область зимовок от Индии до Индокитая, Филиппин и Индонезии, а также ближний мигрант - сибирская чечевица с маньчжурско-корейско-японской областью зимовки.
Во вторую группу включены большая горлица, обыкновенная чечевица, рыжая и желтобровая овсянки, пеночка-зарничка и синехвостка. Для них общим признаком является китайская зимовка. Третий кластер объединяет горную трясогузку и зеленую пеночку, зимовочная область которых включает пакистано-индийскую и индокитайскую зимовки. Еще одна группа включает пестрого дрозда и глухую кукушку, зимовочный ареал которых охватывает филиппино-малакко-индонезийскую зимовку, а также ближнего мигранта - сибирскую завирушку с маньчжурско-корейско-японской зимовкой. В четвертый кластер входят малая мухоловка и пятнистый конек с широкой областью зимнего пребывания от Индии до Индокитая.
В последнюю группу дальних мигрантов включены обыкновенная кукушка с африканской областью зимовки и вьюрок, ведущий номадный образ жизни и улетающий зимовать в разные годы от Маньчжурии до Южного Китая. К ним тяготеет зимующий в Индии, Бирме, Южном Китае и Индокитае дубровник, численность которого повсеместно, в том числе и в горах юга Восточной Сибири, катастрофически снизилась в последние 10Ц15 лет (рис. 5).
Ближние мигранты распределились достаточно независимо: крапивник, белошапочная овсянка и серый снегирь сформировали самостоятельные кластеры, сибирская чечевица и сибирская завирушка вошли в разные кластеры с участием дальних мигрантов, а чиж оказался включенным в группу с синицами.
Максимальная доля видов, снижающих обилие, относится к периферийной и погранично-ареальной группам (35,7 и 42,9 %, соответственно) (табл. 7).
В результате необходимо указать, что выявленная классификационная схема не отражает ареалогические, биотопические, трофические или систематические особенности включенных в анализ видов. Максимальное сходство долговременных изменений обилия виды птиц имеют по местам преимущественных зимовок.
Таблица 7
Состав групп видов птиц ключевого участка с различными трендами долговременных изменений гнездового обилия в 1984Ц2007 гг. (число видов)
Типа группировки |
Обилие птиц |
|||||||
возрастает |
снижается |
стабильное |
||||||
абс. |
% |
абс. |
% |
абс. |
% |
|||
Район зимовки |
||||||||
Оседлые |
3 |
20,0 |
3 |
20,0 |
9 |
60,0 |
||
Ближние мигранты |
1 |
14,3 |
2 |
28,6 |
4 |
57,1 |
||
Дальние мигранты |
4 |
12,1 |
11 |
33,3 |
18 |
54,6 |
||
Местоположение района исследований в ареале вида |
||||||||
Центр ареала |
4 |
11,8 |
8 |
23,5 |
22 |
64,7 |
||
Периферия ареала |
4 |
28,6 |
5 |
35,7 |
5 |
35,7 |
||
Граница ареала |
0 |
0 |
3 |
42,9 |
4 |
57,1 |
||
Всего: |
8 |
14,5 |
16 |
29,1 |
31 |
56,4 |
||
5.3. Связь долговременных изменений обилия видов птиц с их плотностью в предыдущий год
Такая гипотеза опирается на предположения, высказываемые рядом орнитологов (Соколов, 1999; Михантьев, Селиванова, 2005; Головатин, 2007; Носкова, 2007), что уровень осенней численности вида в предыдущем году отражается на изменениях его обилия в год наблюдений. В то же время многие исследователи указывают на отсутствие такой связи (Паевский, 1981, 1985, 2008; Данилов и др., 1984; Зимин, 1988; DeSante, 1990; Рябицев, 1993; Лихачев, 2002; Преображенская, 2003).
Непосредственная оценка связи видовой плотности пернатых в том или ином выделе в первую и вторую половину лета с уровнем обилия в гнездовой сезон следующего года для фоновых видов не выявила статистически значимого уровня такой корреляции. Статистически существенная положительная связь гнездовой плотности оседлого вида в текущем году с обилием в предыдущий летний сезон обнаружена только для буроголовой гаички (rS = +0,53, p < 0,01) и белокрылого клеста (rS = +0,41, p < 0,05).
Положительные статистически значимые связи выявлены между зимней плотностью вида на ключевом участке и его обилием в следующий гнездовой сезон для обыкновенного поползня (rS = +0,65, p < 0,001), желтоголового королька (rS = +0,69, p < 0,001), кедровки (rS = +0,60, p < 0,01), обыкновенной пищухи (rS = +0,59, p < 0,01) и белокрылого клеста (rS = +0,43, p < 0,05). Уровень зимней плотности населения этих видов определяет их летнее обилие в следующий сезон размножения и свидетельствует об отсутствии значительного перераспределения в предгнездовой период.
Следовательно, не выявлены статистически существенные связи изменений уровня гнездового обилия видов птиц с их летней плотностью в предыдущий год наблюдений, но для некоторых зимующих видов прослеживается зависимость гнездовой плотности от их зимнего обилия.
5.4. Влияние абиотических факторов на динамику обилия видов птиц в гнездовой период
Анализ долговременных изменений гнездовой плотности фоновых видов птиц на ключевом участке и его отдельных выделах выявил влияние межгодовых отличий уровня весенней теплообеспеченности на распределение видов в местообитаниях с отличающимся гидротермическим режимом. В качестве индексов теплообеспеченности использованы суммы активных температур выше 0? С, выше +5? С и выше +10? С за апрель, май и июнь (рассчитаны по данным метеостанции Давша на побережье оз. Байкал).
Для гнездового обилия птиц прослеживается позитивная или негативная связь с показателями теплообеспеченности весеннего сезона. Эти зависимости, как правило, не меняют своего знака в разных выделах (побережье оз. Байкал, нижняя и верхняя части горно-лесного пояса и подгольцово-субальпийский пояс). Следует отметить в целом для всего ключевого участка достоверную позитивную связь (р < 0,05) с показателями теплообеспеченности для синехвостки, таежной мухоловки, пеночки-зарнички, а негативную - для желтобровой овсянки и горной трясогузки, т.е. в первой группе в годы с ранней и теплой весной гнездовая плотность возрастала, а во второй - снижалась.
На предгорном выделе соотношение числа видов с положительными и отрицательными связями обилия с весенней теплообеспеченностью составило 8:9, что близко к соотношению в прибрежно-равнинном выделе - 5:5. В горно-лесном выделе это соотношение сдвинуто в пользу видов с положительными связями (10:4), а в подгольцово-субальпийском преобладают отрицательные статистически значимые связи (2:7). В целом по ключевому участку статистически достоверные связи гнездового обилия с параметрами теплообеспеченности весеннего периода зарегистрированы для 36 видов, в том числе положительные - у 16 и отрицательные - у 14 видов. Для 6 видов отмечены противоположные типы связей в различных высотно-поясных выделах.
Например, в годы с поздней холодной весной увеличивалось обилие желтобровой, седоголовой, белошапочной и рыжей овсянок, горной трясогузки, пятнистого конька, сибирской завирушки, зеленой пеночки, пеночки-таловки, сибирской мухоловки, вьюрка, сибирской чечевицы и перевозчика, что можно связать с возрастающей в такие сезоны долей лоседания этих видов в результате задержки миграционного потока. В то же время повышению теплообеспеченности июня сопутствует, например, повсеместное снижение обилия корольковой пеночки вследствие пролонгации миграции этого поздно прилетающего вида.
Для ряда видов птиц на разных участках наблюдений связь с теплообеспеченностью может изменяться на противоположную. Например, для корольковой пеночки в верхней части горно-лесного пояса корреляция с суммами активных температур положительная, а на прибрежно-равнинном участке - отрицательная, у певчего дрозда в горно-лесном поясе связь положительная, а в предгорном - отрицательная. Это определяется различной степенью продвижения основной части особей в верхние высотные пояса в годы с отличающимися метеоусловиями.
У некоторых видов как среди мигрантов (обыкновенная кукушка, горный конек, обыкновенная чечевица, бурая пеночка, крапивник), так и среди зимующих птиц (рябчик, обыкновенный поползень, московка, длиннохвостая синица, щур, обыкновенная пищуха) достоверные корреляции обилия с параметрами теплообеспеченности в весенний период не выявлены.
Для отдельных видов направленность статистически значимых связей может отличаться в одних и тех же высотно-поясных выделах в различных речных долинах (сибирский жулан, обыкновенная чечевица, пятнистый конек, буроголовая гаичка, длиннохвостая синица). Следовательно, под влиянием абиотических (метеорологических) факторов реализуется избирательное заселение птицами выделов не только в различных высотных поясах, но и в долинах соседних рек, условия обитания в которых могут отличаться при сходных параметрах весенней теплообеспеченности.
Многими исследователями (Соколов, 1999; Sokolov et al., 2000; Аськеев, Аськеев, 2002; Артемьев, 2006 и др.) указывается значимость для изменения гнездового обилия птиц гидротермических условий - температуры и осадков в предгнездовой сезон. Статистически значимые корреляции долговременных изменений видового обилия птиц с показателями среднемесячных температур и месячными суммами осадков обнаруживаются для весны и начала лета (апрель-июнь) в год выполнения учета. Характер связей (положительные или отрицательные связи) не изменяется в различных высотно-поясных выделах. Такой же анализ, выполненный для различных участков соседних речных долин, привел к сходным результатам.
Для некоторых видов выявлены такие зависимости и для весенне-летних месяцев предшествующего года. При этом предполагается, что гидротермические параметры местообитания (температура и осадки) в весенне-летний период предыдущего года через трансформацию обилия доступных кормов влияют на видовую плотность птиц в текущем году, в первую очередь через воздействие на изменение численности беспозвоночных животных (Ольшванг, 1992).
Таким образом, метеорологические параметры весны и начала лета (теплообеспеченность и осадки), определяющие в горных условиях гнездопригодность и состояние кормовых ресурсов местообитаний, обуславливают формирование гнездового населения мигрирующих и оседлых видов птиц.
5.5. Влияние фенологических сроков весны на формирование населения гнездящихся птиц
Весенняя погода определяет весь ход фенологического развития природы, в том числе как прямо, так и опосредованно воздействует на сроки и динамику многих биологических явлений в годовом цикле птиц (Данилов, 1966; Jarvinen, 1989; Рябицев, 1993). Ход апрельских и майских температур воздуха отражает общий процесс весеннего потепления, наступление сроков весенних субсезонов, которые в свою очередь могут выступать в качестве индикаторов состояния природных комплексов, их готовности обеспечить птиц доступными кормовыми и гнездопригодными ресурсами.
Использовано деление фенологического года на сезоны и субсезоны, установленное для Баргузинского хребта К.П. Филоновым (1978). Весенний сезон подразделяется на четыре субсезона: снежная, пестрая, голая и зеленая весна. Дополнительно проанализированы такие феноиндикаторы (явления, регулярно совпадающие с началом или окончанием весенних субсезонов) как асроки начала уменьшения высоты снежного покрова и полного схода снега на открытых местах на побережье оз. Байкал. В качестве модельных объектов были выбраны 12 видов птиц с максимальным среднемноголетним обилием: 5 - оседлых и 7 - перелетных (табл. 8).
Обнаружено влияние межгодовых отличий сроков наступления весенних субсезонов и дат регистрации выбранных феноиндикаторов на распределение обилия птиц между выделами. Для таких оседлых видов как рябчик и буроголовая гаичка выявлено повышение гнездовой плотности в годы с более ранними сроками наступления весны. Связи обилия московки с фенологическими сроками весны имеют противоположный характер, ее плотность во многих выделах выше в годы с поздней весной. Изменения плотности гнездования обыкновенного поползня и пестрого дятла достоверно не связаны со сроками наступления весенних субсезонов. В целом зимующие виды имеют значительно меньшие связи гнездового обилия со сроками наступления весенних субсезонов, которые лишь в незначительной степени влияют на формирование населения.
Мигрирующие виды птиц в большей степени связаны с ходом весенних фенологических процессов. Для всех включенных в анализ перелетных видов, за исключением нерегулярно зимующего чижа, характерно повышение гнездовой плотности в годы с более ранними сроками прохождения весенних фенофаз. В то же время для горной трясогузки и пятнистого конька в различных высотных выделах отмечаются разнонаправленные связи обилия со сроками наступления пестрой и голой весны, что определяется высотно-поясными и междолинными перераспределениями гнездовой плотности. Для чижа более характерно повышение гнездовой плотности в годы с поздней весной, что, возможно, определяется недолетом части мигрирующей через район исследований популяции до лежащих севернее территорий и ее лоседанием для гнездования в южной части гнездового ареала.
В годы с холодной поздней весной обилие птиц в высокогорьях обычно ниже, чем в годы с ранней и теплой весной. Кроме того, в отдельные, наиболее теплые годы в верхних поясах можно наблюдать виды птиц, которые в обычные годы там не встречаются. Например, летом 2005 г. в гольцово-альпийском поясе Баргузинского хребта зарегистрировано массовое гнездование белошапочных овсянок, которые в обычные годы не отмечались выше предгорий. а
Таблица 8
Корреляционные связи гнездового обилия птиц со сроками начала весенних субсезонов
и весенними феноиндикаторами (*p <0.05, **p <0.01)
Вид |
Фенофазы весны |
Снежный покров |
||||
снежная |
пестрая |
голая |
зеленая |
начало схода |
сход |
|
Обыкновенный поползень |
пр-б (+)* |
пр-б (+)*, пг-б (+)* |
||||
Московка |
ку-д (+)* |
ку-б (+)**, ра-б (+)** пг-б (+)* |
||||
Буроголовая гаичка |
пг-д (-)* |
гл-д (-)* |
||||
Корольковая пеночка |
пр-б (-)*, пр-е (-)* пг-д (-)* |
ку-д (-)*, пр-б (-)* |
||||
Синехвостка |
ку (-)*, пр-е (-)* пг-е (-)* |
пг-б (+)*, ку-е (-)* пр-е (-)** |
ку (-)*, ку-б (-)* рм-б (-)*, пг-е (-)* |
ку-е (-)*, пр-е (-)* |
пр-е (-)* гл-е (-)* пг-д (-)* |
|
Пятнистый конек |
пг-б (+)* |
ку-б (+)* пр-б (+)** ра-б (+)* |
ку-б (+)*, пр-б (+)* |
пр-б (+)* ку-е (-)* |
||
Чиж |
ку-д (+)*, пс-д (+)** |
|||||
Зеленая пеночка |
пр-б (-)* |
пг-е (+)* |
ку-б (-)*, пг-б (-)* |
ку-е(-)** |
||
Пеночка-зарничка |
ку (-)*, пг-е (-)* пр-д (-)* |
пр-б (-)* |
пг-б (+)* |
|||
Рябчик |
ку-д (-)* |
гл-е(+)** |
ку-д (-)**, пг-д (-)* |
пг-д (-)* |
||
Горная трясогузка |
ра-б (+)** пр-д (-)* |
пр-д(-)** |
||||
Пестрый дятел |
ку-д (-)*, пр-д (-)* пг-д (-)* |
пг-е (+)* |
||||
Примечания: (+) - корреляция положительная; (-) - корреляция отрицательная. Наименования выделов: ПР - прибрежно-равнинный, РА - равнинный, РМ - равнинно-моренный, ПГ - предгорный, ГЛ - горно-лесной, ПС - подгольцово-субальпийский, КУ - ключевой участок; ЦБ - долина р. Большая, ЦЕ - долина р. Езовка, ЦД - долина р. Давша.
Исходя из наших многолетних наблюдений можно заключить, что сроки развития фенологических процессов в природных комплексах влияют на формирование местного населения за счет изменения количества обосновавшихся в районе гнездования особей, вызывая разнонаправленные изменения обилия в различных высотно-поясных выделах и в соседних речных долинах, отличающихся по своим экологическим условиям.
5.6. Долговременные изменения сроков прилета птиц и их влияние на формирование гнездового населения
Изменения дат весеннего прилета птиц, в последние десятилетия регистрируются в различных регионах Северного полушария, как в Европе (Jarvinen, 1995; Moss, 1996, 1998; Bergmann, 1998; Loxton et al., 1998; Sokolov et al., 1998; Соколов и др., 1999; Gilyazov, Sparks, 2002; Соколов, 2006 и др.), так и в Северной Азии (Кучин, 1996, 2001; Пасхальный, 2002).
Межгодовые отклонения сроков весеннего прилета многих видов птиц достаточно тесно взаимосвязаны с весенними температурами (Михеев, 1964; Гаврилов, 1979; Гаврин и др., 1980; Кривенко и др., 1980; Гордиенко, 1983; Сема, 1989; Alerstam, 1990; Huin, Sparks, 1998; Sokolov et al., 1998; Sokolov, 2000; Sparks, Loxton, 2002; Соколов, 2006, 2007 и др.).
На Баргузинском хребте изменения климата выразились в потеплении весенних и летних месяцев и, как следствие, в увеличении среднегодовой температуры воздуха, удлинении безморозного периода, более раннем освобождении ото льда акватории озера Байкал и изменении других параметров (Ананин и др., 2001; Ананин, Ананина, 2002). В динамике среднегодовой температуры и средней температуры воздуха отдельных весенних месяцев (март, апрель, май) выявлены положительные тренды.
Анализ долговременных трендов дат первой весенней регистрации для 54 видов птиц показал, что для 28 видов (51,8 %) отмечается тенденция к более раннему прилету. В 1984Ц2007 гг. для 6 из них (полевой лунь, чибис, воронок, полевой жаворонок, черноголовый чекан, юрок) зарегистрирован прилет на 10 и более дней раньше, чем 30Ц50 лет назад. Еще для 8 видов (черныш, большая горлица, деревенская ласточка, горная трясогузка, сибирский жулан, обыкновенный скворец, сибирская завирушка, краснозобый дрозд) опережение сроков первой регистрации составило 7-9 дней. Для 15 видов (27,8%) зарегистрировано более позднее появление весной. Половину позднее прилетающих видов составили околоводные птицы. Сроки весеннего прилета еще 11 видов птиц (20,4%) достоверно не изменились (табл. 9).
Более ранние весенние регистрации связываются с долговременными тенденциями потепления климата. Для большинства видов с отрицательным трендом сроков прилета выявлена значимая негативная связь (p < 0.05) с температурами весенних месяцев (или соответствующих декад), в которые регистрируется их прилет. Не обнаружены связи изменений сроков весеннего прилета с местами зимовок и протяженностью миграционного пути.
Таблица 9
Соотношение числа видов, проявивших тенденцию к более раннему и позднему прилету (Баргузинский хребет, 1938 - 2007 гг.)
|
Отряд |
Число видов |
|||
всего |
прилетающих раньше |
прилетающих позже |
прилетающих в те же сроки |
|
Anseriformes |
5 |
|
4 (80.0 %) |
1 (20.0 %) |
Falconiformes |
6 |
1 (16,7 %) |
4 (66.7) |
1 (16.7) |
Gruiformes |
1 |
1 |
|
|
Charadriiformes |
9 |
3 (33.3) |
2 (22.2) |
4 (44.4) |
Columbiformes |
1 |
1 |
|
|
Cuculiformes |
2 |
2 |
|
|
Coraciiformes |
1 |
1 |
|
|
Passeriformes |
29 |
19 (65.5) |
5 (17.2) |
5 (17.2) |
Итого |
54 |
28 (51.9) |
15 (27.8) |
11 (20.4) |
При более раннем прилете обыкновенной кукушки отмечено увеличение ее плотности на прибрежно-равнинном участке и снижение - на более удаленном от побережья оз. Байкал равнинном. Повышение гнездовой плотности при более раннем прилете отмечено для глухой кукушки, пятнистого конька, пеночки-зарнички, малой мухоловки и вьюрка, а снижение - для горного конька, горной трясогузки, соловья-красношейки, белошапочной и желтобровой овсянок. Для трех включенных в анализ видов (синехвостка, краснозобый дрозд и обыкновенная чечевица) статистически значимая связь гнездового обилия со сроками прилета не обнаружена.
Таким образом, анализ долговременных изменений сроков прилета птиц, проведенный впервые в горных условиях Восточной Сибири, позволил выявить основные тренды их изменений, которые влияют на процессы формирования локального обилия видов. Глобальные изменения климата в регионе через сдвиги сроков прилета птиц могут оказывать воздействие на формирование местного населения и сопровождаться направленными трансформациями гнездовой плотности птиц.
Глава 6. Роль экологических факторов в долговременной динамикЕ зимнего обилия птиц
6.1. Динамика обилия фоновых видов птиц в зимний период
Общий уровень плотности зимующего населения птиц определяется в основном флуктуациями обилия многочисленных оседлых и нерегулярно зимующих видов. Наиболее резкие колебания обилия характерны для инвазионных, нерегулярно зимующих видов - белокрылого клеста и обыкновенной чечетки (рис. 7). Чаще годовые различия достигали 10-30-кратной величины (пестрый и трехпалый дятлы, желна, рябчик, кукша, кедровка, длиннохвостая синица, пищуха и желтоголовый королек).
Таблица 10
Корреляционные связи гнездовой плотности птиц на высотно-поясных выделах со сроками прилета
на побережье оз. Байкал (*p <0.05, ** p <0.01)
Вид |
Высотно-поясные выделы |
|||||
Весь участок |
Прибрежно-равнинный |
Предгорный |
Горно-лесной |
Подгольцово-субальпийский |
Гольцово-альпийский |
|
Обыкновенная кукушка |
-0,11 |
0,04 |
-0,05 |
0,13 |
-0,05 |
-0,13 |
Глухая кукушка |
-0,18 |
-0,03 |
-0,10 |
-0,15 |
-0,12 |
-0,18 |
Пятнистый конек |
-0,23 |
-0,44* |
0,19 |
0,14 |
0,09 |
0,15 |
Горный конек |
|
|
|
|
0,47* |
0,33 |
Горная трясогузка |
0,44** |
0,23 |
0,27 |
0,33* |
0,19 |
-0,22 |
Пеночка-зарничка |
0,14 |
0,02 |
0,06 |
0,20 |
-0,48* |
-0,68** |
Корольковая пеночка |
-0,14 |
0,14 |
0,25 |
-0,37* |
0,59* |
|
Малая мухоловка |
-0,59** |
-0,22 |
-0,18 |
-0,06 |
|
|
Соловей-красношейка |
0,46* |
|
0,19 |
0,17 |
0,13 |
-0,04 |
Синехвостка |
0,06 |
0,17 |
0,15 |
-0,04 |
0,17 |
|
Краснозобый дрозд |
-0,13 |
|
|
-0,19 |
0,04 |
0,11 |
Вьюрок |
-0,11 |
0,12 |
0,10 |
-0,09 |
-0,12 |
-0,52* |
Обыкновенная чечевица |
-0,05 |
|
0,17 |
-0,22 |
-0,08 |
0,07 |
Белошапочная овсянка |
0,09 |
0,13 |
|
-0,07 |
|
|
Желтобровая овсянка |
0,25 |
0,46* |
-0,15 |
0,00 |
|
|
Примечания: пропуск данных - вид регулярно в данном выделе не обитает.
И только у трех наиболее массовых видов (буроголовая гаичка, московка и обыкновенный поползень) обилие изменялось в минимальных пределах (от 4,5 до 5,5 крат).
Наиболее похожими на периодические оказались колебания зимнего обилия щуров и пестрых дятлов. Подъемы их плотности происходили с интервалом в 9 и 7 лет, соответственно. Для обыкновенной пищухи такие пики зафиксированы достаточно регулярно, через 4Ц5 лет. Для буроголовой гаички и обыкновенного поползня максимумы обилия отмечались с высокой степенью синхронности, через 2Ц3 года. В конце периода наблюдений для них зарегистрированы пропуски пика численности, когда интервал между подъемами обилия составил 5 лет (рис. 7). Максимумы обилия обыкновенной чечетки, белокрылого клеста, желтоголового королька, кедровки, кукши, а также трехпалого дятла и рябчика обычно повторялись хаотично, без определенной периодичности, либо через 2Ц4 года.
|
|
|
|
|
|
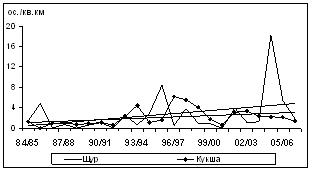
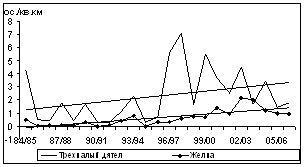
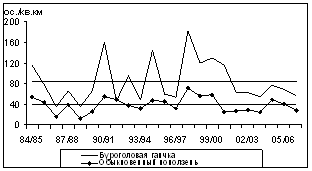
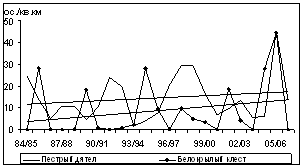
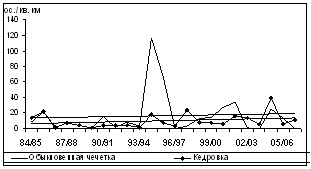

Рис. 7. Изменения обилия зимующих видов птиц ключевого участка в субсезон морозной зимы (1984/85Ц2006/07 гг., январь-февраль, особ./км2)
У 3 видов птиц из 20, регулярно встречавшихся на зимних учетах, за 23-летний период наблюдений отмечено повышение обилия (щур, кукша и желна). Еще у 3 видов зарегистрирован положительный тренд в отдельных высотно-поясных выделах (на прибрежно-равнинном - у обыкновенной пищухи, на горно-лесном - у рябчика, и на обоих этих выделах - у трехпалого дятла). У 14 видов не обнаружено достоверных тенденций изменений. Отрицательный тренд многолетних изменений обилия не выявлен.
Синхронность колебаний не является строгой закономерностью для представителей зимнего населения птиц региона, как это было отмечено для синичьих стай в Подмосковных лесах (Боголюбов, Преображенская, 1989). Изменения обилия были параллельными во времени лишь между несколькими видами разных групп и не совпадали у других видов. Дендрограмма сходства динамики плотности зимующих на ключевом участке видов птиц выявляет низкий уровень различий между долговременными изменениями обилия (рис. 8). Наиболее близки объединенные в один кластер малочисленные и нерегулярно зимующие виды (чиж, сибирская чечевица, серый снегирь, обыкновенный клест, седой дятел и желна).
Вторую группу составляют насекомоядные виды, обилие которых не связано с урожайностью древесных семян: кукша, трехпалый дятел и обыкновенная пищуха. Отдельные, но достаточно связанные между собой кластеры последовательно формируют щур, желтоголовый королек и рябчик. Следующая группа включает кедровку и белокрылого клеста, зимняя плотность которых значительно колеблется в зависимости от урожая семян кедра и лиственницы. Еще один самостоятельный кластер формируют пестрый дятел и длиннохвостая синица, у которых уровень плотности достаточно изменчив, но не связан с урожайностью кедра и лиственницы. Обыкновенная чечетка, обилие которой связано с урожайностью берез, образует отдельный кластер.
Совершенно самостоятельную группу формируют пара достаточно близких по характеру долговременных изменений плотности населения видов (обыкновенный поползень и московка), а также наиболее удаленная от них буроголовая гаичка (рис. 8).
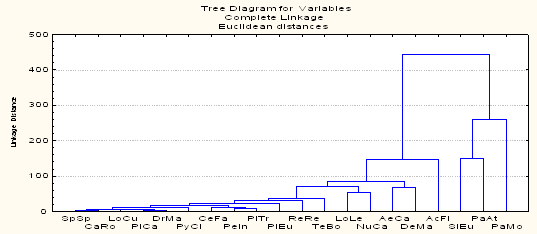
Рис. 8. Дендрограмма сходства межгодовых изменений обилия зимующих видов птиц на ключевом участке (1984/85Ц2006/07 гг.): обозначения видов птиц даны по первым двум буквам латинского названия рода и вида.
Следовательно, общий уровень плотности зимнего населения птиц определяется в основном флуктуациями обилия немногих наиболее многочисленных оседлых и нерегулярно зимующих видов. Годы депрессий характеризуются низкой плотностью буроголовой гаички, московки, обыкновенного поползня и отсутствием белокрылых клестов и обыкновенных чечеток, а годы подъемов - обратной картиной. Для ряда видов в период исследований выявлены периодические подъемы обилия, а также положительный тренд долговременных изменений плотности.
6.2. Влияние кормообеспченности на обилие зимующих видов птиц
Разнообразие факторов среды, воздействующих на зимующих в регионе птиц, вызывает появление у них различных адаптаций, помогающих пережить зиму. Наиболее значимым фактором оказывается питание. В зимнем питании большинства видов птиц Восточной Сибири важную роль играет массовый и легкодоступный растительный корм. Поиск такого корма связан с меньшими затратами энергии, что в условиях суровых зим биологически более целесообразно (Зонов, 1982).
Главная особенность приспособления птиц к условиям существования зимой - выбор кормового объекта, наиболее обеспечивающего организм энергетическими ресурсами при относительно экономном расходе энергии на добывание кормов. Адаптивное значение поедания зимой семян насекомоядными птицами определяется тем, что это массовый, территориально сконцентрированный и легкодоступный корм. В условиях суровых зим добывание семян менее трудоемко, чем скрытых в убежищах насекомых.
Для многих зимующих птиц существенное значение имеет урожай древесных семян. Как правило, высокая численность обыкновенных клестов является следствием обильного плодоношения ели, белокрылых клестов - урожая семян лиственницы, чечеток - семян березы, пестрых дятлов - сосны и в какой-то степени ели и лиственницы, кедровок - сибирского кедра (Новиков, 1956; Филонов, 1960, 1961, 1962а-б; Формозов, 1976; Мальчевский, Пукинский, 1983 и др.).
Из других регионов известно, что осеннее обеспечение семенами древесных растений может во взаимодействии с зимней погодой управлять весенним обилием птиц (Wesolowski, Tomialojc, 1997), например, обыкновенного поползня (Enoksson, Nilsson, 1983; Nilsson, 1987; Matthysen, 1989), пестрого дятла (Glutz von Blotzheim, 1980), синиц (Svensson, 1981; Newton, 1994) и чижа (Eriksson, 1970).
В результате анализа многолетней динамики зимнего обилия 16 оседлых видов птиц выявлено, что 3 вида (рябчик, белокрылый клест и кукша) в гнездовой и зимний периоды предпочитают прибрежно-равнинный участок, 8 видов летом и 6 видов зимой - предгорный участок, а горно-лесной участок - 4 вида в период гнездования и 6 видов зимой.
Для 12 видов (75,0 %) область преференции (предпочтения) в гнездовой и зимний периоды в среднем не изменялась, для московки, желны и седого дятла (18,7 %) она повышалась, предпочтение в зимний период отдавалось вышележащему высотному поясу, а для длиннохвостой синицы (6,3 %) - снижалась от предгорного участка к прибрежно-равнинному. Область преференции рассчитана как доля сезонов, в которые участок имел максимальное значение обилия вида среди выделов лесного пояса.
Отклонения от среднемноголетних территорий зимнего предпочтения для семеноядных видов обусловлены в первую очередь перераспределением участков с обильным плодоношением основных кормов. Так, например, изменение обилия зимующих щуров в различных высотно-поясных выделах связано со значительным урожаем семян лиственницы на прибрежно-равнинном или предгорном участках, либо с высоким урожаем семян ели и пихты в горно-лесном поясе. Для пестрого дятла такое перераспределение определяется размещением урожайных участков сосны или лиственницы, а для трехпалого дятла и желны - наличием свежих участков гарей с массовым усыханием поврежденных огнем хвойных насаждений (табл. 11). Перераспределение оседлых видов между высотными выделами происходит в процессе позднелетних и осенних кочевок.
Таким образом, нашими многолетними наблюдениями подтверждено, что в основе выбора участков зимнего обитания птиц лежит поисковое поведение, направленное на выявление территорий с доступной и достаточной кормовой базой. Наличие убежищ и конкурентные отношения с видами со сходными кормовыми предпочтениями имеют дополнительное значение.
6.3. Связь долговременных изменений зимнего обилия видов птиц с их плотностью в предыдущий год
Статистически значимая положительная связь между плотностью гнездящихся птиц в I и II половину лета и их обилием в следующую зиму не прослежена, что доказывает высокий уровень послегнездовых перемещений у группы оседлых видов. а
Наибольшая тенденция к позднелетним и осенним кочевкам выявляется для щура, пестрого дятла, длиннохвостой синицы и московки (rS = Ц0,33 Ц0,14, n. s.). Для трехпалого дятла, желны и обыкновенной пищухи проявляется тенденция к низкому уровню послегнездовых перемещений (rS = +0,38 - +0,29, n. s.).
Статистически значимая положительная связь зимней плотности птиц с видовым обилием в предыдущий зимний сезон обнаружена для оседлых видов: желны (rS = +0,67, p < 0,001) и кукши (rS = +0,44, p < 0,05), а также для нерегулярно зимующих сибирской чечевицы (rS = +0,45, p < 0,05) и серого снегиря (rS = +0,43, p < 0,05).
Следовательно, для зимующих на Баргузинском хребте видов птиц не прослеживается тесная связь зимнего обилия с летней плотностью предыдущего года, что определяется высоким уровнем осенних кочевок.
Таблица 11
Изменение зоны преференции (предпочтения) оседлых видов птиц в гнездовой и зимний периоды
на ключевом участке (1984-2007 гг.)
Вид |
Плотность, ос./км2 |
Область преференции, % |
||||||||||
Гнездовой период (1 июня - 15 июля) |
Морозная зима (январь - февраль) |
Гнездовой период (1 июня - 15 июля) |
Морозная зима (январь - февраль) |
|||||||||
прибрежно-равнинный |
предгорный |
горно-лесной |
прибрежно-равнинный |
предгорный |
горно-лесной |
прибрежно-равнинный |
предгорный |
горно-лесной |
прибрежно-равнинный |
предгорный |
горно-лесной |
|
Поползень |
21,7 |
34,8 |
23,1 |
28,7 |
45,1 |
42,7 |
8,3 |
87,5 |
4,2 |
0,0 |
54,2 |
45,8 |
Буроголовая гаичка |
15,7 |
18,5 |
24,6 |
57,6 |
80,8 |
108,8 |
20,8 |
20,8 |
58,3 |
0,0 |
25,0 |
75,0 |
Московка |
20,4 |
28,2 |
11,9 |
15,4 |
74,0 |
102,0 |
16,7 |
75,0 |
8,3 |
0,0 |
25,0 |
75,0 |
Рябчик |
12,2 |
11,1 |
5,2 |
7,9 |
4,1 |
7,9 |
60,9 |
34,8 |
4,3 |
41,7 |
16,7 |
41,7 |
Кедровка |
7,7 |
8,5 |
6,7 |
7,9 |
11,9 |
10,6 |
34,8 |
34,8 |
30,4 |
12,5 |
37,5 |
50,0 |
Пестрый дятел |
6,5 |
12,1 |
1,3 |
14,1 |
23,7 |
5,5 |
20,8 |
79,2 |
0,0 |
4,2 |
95,8 |
0,0 |
Белокрылый клест |
8,8 |
5,3 |
4,4 |
13,3 |
12,6 |
3,7 |
56,5 |
17,4 |
26,1 |
61,1 |
22,2 |
16,7 |
Кукша |
5,3 |
4,6 |
2,5 |
2,6 |
1,9 |
1,5 |
56,5 |
34,8 |
8,7 |
65,2 |
21,7 |
13,0 |
Обыкновенная пищуха |
2,4 |
5,2 |
1,0 |
1,7 |
2,6 |
2,1 |
14,3 |
81,0 |
4,8 |
26,3 |
36,8 |
36,8 |
Желтоголовый королек |
0,9 |
4,8 |
2,4 |
0,2 |
5,1 |
3,1 |
0,0 |
75,0 |
25,0 |
0,0 |
66,7 |
33,3 |
Трехпалый дятел |
1,9 |
3,0 |
3,4 |
1,5 |
2,6 |
2,9 |
13,6 |
36,4 |
50,0 |
16,7 |
33,3 |
50,0 |
Длиннохвостая синица |
1,4 |
2,3 |
1,1 |
16,4 |
13,0 |
7,4 |
27,8 |
55,6 |
16,7 |
59,1 |
31,8 |
9,1 |
Оляпка |
0,0 |
0,2 |
2,7 |
0,0 |
0,0 |
0,2 |
0,0 |
6,3 |
93,8 |
0,0 |
0,0 |
100,0 |
Щур |
0,0 |
0,0 |
2,5 |
2,6 |
2,2 |
2,9 |
0,0 |
0,0 |
100,0 |
28,6 |
19,0 |
52,4 |
Желна |
0,5 |
1,3 |
0,3 |
0,5 |
0,5 |
1,1 |
20,8 |
75,0 |
4,2 |
31,8 |
22,7 |
45,5 |
Седой дятел |
0,0 |
0,07 |
0,01 |
0,02 |
0,3 |
0,2 |
0,0 |
50,0 |
50,0 |
14,3 |
42,9 |
42,9 |
6.4. Значение абиотических факторов в динамике обилия зимующих видов птиц
Многолетние колебания обилия отдельных видов зимующих птиц определяются не только наличием кормов, но и погодно-климатическими условиями в период предзимних кочевок и самой зимовки (Филонов, 1961, 1962; Eriksson, 1970; Rotenberry et al., 1979; Боголюбов, Преображенская, 1989). Имеются многочисленные исследования, указывающие на то, что численность оседлых птиц в Западной Европе и Фенноскандии строго зависит от суровости предыдущей зимы (von Haartman, 1971; van Balen, 1980; Cawthorne, Marchant, 1980; Kallander, Karlsson, 1981; Svensson, 1981; Hilden, 1982; Nilsson, 1986, 1987; Wesolowski, Tomialojc, 1997 и др.). Сильные морозы в Европе могут вызвать катастрофические последствия для воробьиных, что связано со значением сезонной настройки системы терморегуляции у птиц (Шилов, 1968). Постоянно низкая температура в районах Восточной Сибири не оказывает на птиц столь губительного действия, как похолодания в более умеренных регионах Европы (Зонов, 1982). Адаптации птиц горных систем юга Восточной Сибири к отрицательным условиям погоды осуществляются, в том числе, активным выбором склонов, лучше освещенных солнцем, и таких участков, где действие ветра и снегопадов минимально. Теневые участки гор птицы зимой обычно избегают.
Анализ влияния метеорологических характеристик зимних месяцев (сумма осадков и высота снежного покрова, среднесуточные, максимальные и минимальные температуры воздуха октября-февраля) на обилие зимующих птиц в субсезон морозной зимы (январь-февраль) показал, что из 30 видов лишь у половины выявлены статистически значимые корреляционные связи обилия с одним, двумя или очень редко тремя метеорологическими параметрами (табл. 12).
Тенденция увеличения видовой плотности в относительно теплые зимы прослеживается у белокрылого клеста, сойки и сибирской чечевицы, а в холодные зимы отмечается повышение обилия буроголовой гаички, московки, обыкновенного поползня и длиннохвостой синицы в предгорной части, для щура - в горно-лесном выделе, а для трехпалого дятла - на всем ключевом участке. Следует отметить положительные связи среднемесячных температур октября (субсезон глубокой осени) с плотностью населения московки в горно-лесном поясе и сойки - в прибрежно-равнинном выделе, а также снижение обилия длиннохвостых синиц в предгорном выделе в годы с теплым октябрем.
Увеличение количества осадков в январе, сопутствующее, как правило, более высоким температурам, сопровождалось повышением обилия белокрылого клеста в оптимальном для вида (зона предпочтения или преференции) прибрежно-равнинном выделе, обыкновенной пищухи в субоптимальных местообитаниях прибрежно-равнинного и горно-лесного выделов, сибирской чечевицы - в горно-лесном поясе, а щура и кукши - на всем ключевом участке. Такой характер связи определяется, вероятно, снижением уровня зимней смертности.
Таблица 12
Связи обилия зимующих птиц Баргузинского хребта с зимними температурами и осадками (1984/85Ц2006/07 гг.)
Вид |
Выделы |
|||
прибрежно-равнинный |
предгорный |
горно-лесной |
ключевой участок |
|
Parusmontanus |
|
ЦТ3* |
|
|
Parusater |
ЦО4* |
ЦТ4* |
+Т0* |
|
Sittaeuropaea |
|
ЦТ3* |
+О2* |
+О2* |
Acanthisflammea |
|
ЦО4* |
ЦО4* |
ЦО4* |
Dendrocoposmajor |
+О2* |
+О2*** |
+О2** |
+О2** |
Aegithaloscaudatus |
|
ЦТ0*, ЦТ1**, ЦТ3** |
|
ЦТ1*, ЦТ3* |
Loxialeucoptera |
+Т1*, +О3* |
+Т1* |
+Т1* |
+Т1* |
Tetrastes bonasia |
+О2** |
|
|
+О2* |
Regulusregulus |
|
ЦО2* |
ЦО2* |
ЦО2** |
Certhiafamiliaris |
+О3** |
ЦО2* |
+О3** |
|
Pinicola enucleator |
|
+О3* |
ЦТ4* |
+О3* |
Picoidestridactylus |
|
ЦТ3* |
ЦТ3** |
ЦТ3** |
Perisoreus infaustus |
|
|
|
+О3* |
Garrulusglandarius |
+Т0**, +Т1* |
|
|
+Т0**, +Т1* |
Loxiacurvirostra |
|
|
+О2* |
+О1*, +О2* |
Carpodacusroseus |
|
|
+Т3*, +О3* |
+Т3*, +О3* |
Примечания. Т - среднемесячная температура; О - сумма осадков за месяц; 0 - октябрь; 1 - ноябрь; 2 - декабрь; 3 - январь; 4 - февраль; + - связь положительная; Ц - связь отрицательная; пропуск данных указывает на отсутствие значимой связи; *p < 0,05, **p < 0,01, ***p < 0,001
При этом следует учитывать, что погодные условия зимы могут иметь корреляционные зависимости, не имеющие характера причинно-следственных связей, с урожайностью основных кормов растительноядных видов птиц, питающихся семенами деревьев и кустарников. Например, высокий урожай рябины предшествует зимам с пониженными температурами января и февраля (p < 0,05). Поэтому у части видов могут выявляться связи с климатическими параметрами зимы, имеющие случайный характер, за счет параллельных корреляций с каким-либо фактором, не включенным в анализ.
Результаты выполненных исследований подтверждают, что в горных условиях юга Восточной Сибири метеорологические показатели зимнего периода не оказывают столь существенного воздействия на уровень обилия большинства зимующих видов птиц, как в различных регионах Европы. Лишь немногие оседлые и нерегулярно зимующие виды выявляют статистически значимый уровень корреляции с зимними температурами и количеством осадков.
Глава 7. Экологические закономерности формирования локального населения птиц в горных условиях
Результаты современных исследований показывают, что распределение гнездящихся птиц в пространстве в первую очередь определяется наличием и доступностью кормовых ресурсов и гнездопригодностью местообитания. Они обусловлены такими параметрами видовой экологической ниши как благоприятный субстрат, архитектурные особенности места обитания (топоархитектура), высотно-климатические условия и комплекс приспособлений к условиям зимовки (Юдкин, 2000, 2009 и др.). При изучении динамики плотности птиц на постоянных участках в горных условиях, где относительно стабильны пригодная для вида топоархитектура и субстрат, наибольшее влияние на распространение и обилие оказывают изменяющиеся параметры - климатические факторы и выживаемость на местах зимовки, поскольку роль изменяющихся факторов намного больше, чем факторов устойчивых (Мончадский, 1958; Пыжьянов, 2006)..
Известно, что существенная часть выживших взрослых особей не возвращается к месту предыдущего гнездования (Соколов, 1991; Высоцкий, 2001). В районах, где условия существования стабильны, птицы проявляют большую верность прежней территории гнездования, чем в районах с резкими колебаниями внешних факторов среды (McNicholl, 1975; Skeel, 1983; Михельсон и др., 1986; Соколов, 1988; Паевский, 2008). В более южных горных системах Палеарктики гнездовой консерватизм выше (Ковшарь, 1979; Бёме, Банин, 2001). В периферийных популяциях некоторых воробьиных птиц выявлено непостоянство местного гнездового населения и его практически полное ежегодное обновление за счет непрерывного притока из зоны регулярного гнездования (Данилов и др., 1984; Шутов, 1986; Зимин, 1988; DeSante, 1990; Рябицев, 1993; Hoffmann, Blows, 1994; Тюлькин, 1995; Рыжановский, 1997; Артемьев, 2006; Thinstad et al., 2006).
В условиях горных систем юга Восточной Сибири уровень возврата ранее гнездившихся взрослых птиц, как правило, также невелик, так как значительная часть видов находятся в регионе на краю или вблизи границы гнездового ареала, поэтому население птиц в регионе практически ежегодно формируется в значительной степени заново из особей-иммигрантов, что подтверждается и данными кольцевания.
С другой стороны, уровень зимней плотности некоторых видов (обыкновенный поползень, желтоголовый королек, кедровка, обыкновенная пищуха и белокрылый клест) в горных условиях определяет их летнее обилие в следующий сезон размножения, что свидетельствует об отсутствии значительного перераспределения этих зимующих видов в предгнездовой период. Обилие буроголовой гаички в текущем году статистически значимо связано ее с гнездовой плотностью в предыдущем году вследствие характерного для вида высокого уровня гнездового консерватизма.
Для определения механизмов формирования местного населения птиц и прогнозирования их локальных изменений обилия необходимо выявление роли внешних и внутрипопуляционных факторов, обуславливающих закономерности этих процессов. Разные виды неодинаково реагируют на складывающуюся ежегодно ситуацию.
Основной группой факторов, оказывающих модифицирующее воздействие на формирование населения птиц, могут быть условия текущей весны, в том числе сроки наступления феноявлений, теплообеспеченность и влагообеспеченность весны и начала лета. Особенно это проявлялось в годы либо с очень ранней и теплой весной, либо при холодной поздней весне.
Выбор оптимальных условий для вида в конкретном году может происходить путем перераспределения между участками в пределах одного высотного пояса (между речными долинами), а также за счет вертикальных перемещений между разными высотными поясами растительности.
В раннюю и теплую весну многие виды гнездятся в верхних высотных поясах Баргузинского хребта с более высокой плотностью, чем в нижележащих, а в холодные, поздние и затяжные весны имеют повышенную плотность в нижних поясах и редки в высокогорной части. Кроме того, в отдельные, наиболее теплые годы, в верхних поясах можно наблюдать гнездование птиц, которые в обычные годы там не встречаются (белошапочная овсянка в подгольцово-субальпийском поясе).
На процессы формирования гнездового населения в горных условиях могут оказывать влияние условия предыдущего года, включающие баланс тепла и влагообеспеченности, составляющий основу для формирования величины биомассы беспозвоночных животных в текущем году. а
Кроме того, существенная роль в процессах формирования населения принадлежит внутрипопуляционным механизмам саморегуляции. В горных условиях региона к их числу могут относиться послегнездовые кочевки оседлых видов, которые в отдельные годы перерастают в массовые миграции. Почти ежегодно наблюдаются массовые перемещения вдоль побережья Байкала, как в северном, так и в южном направлениях, таких видов как московка, буроголовая гаичка, обыкновенный поползень, длиннохвостая синица и кедровка.
Птицы, возвращающиеся с зимовок, совершают в предгнездовой период поисковые перемещения, выявляя территории, наиболее благоприятные для гнездования. Гнездовые поселения птиц формируются весной зачастую до того, как растает снег и еще невозможно оценить или прогнозировать кормность или гнездопригодность местообитаний. В таких случаях велика роль сигнальной информации, которая позволяет птицам делать свой выбор конкретного участка обитания в условиях текущего весенне-летнего периода.
Весеннее поисковое поведение направлено на выявление и занятие гнездовых участков и мест размещения гнезда, которое определяется, как показывают современные исследования, не только непосредственно экологическими факторами, но и наличием предпочитаемых сигнальных характеристик биотопа (McCallum, Gehlbach, 1988; Moller, 1988; Михайлов, 1992). Поэтому для птиц очень значимы релизеры - индикаторы состояния среды обитания в конкретном месте в конкретное время. К числу таких индикаторов (релизеров) следует отнести:
- структуру местообитания, в том числе структуру растительности (топоархитектуру);
- фенологическое состояние развития растительности (существенно зависит от теплообеспеченности, высоты снежного покрова к началу весны, влагообеспеченности для засушливых местообитаний);
- маркеры успешного размножения (или возможности размножения) в предыдущем году, в том числе сохранившиеся старые гнезда;
- наличие в сообществе других особей своего вида, заселивших местообитание ранее, чем находящаяся в поисках места для гнездования особь, высокая значимость рано прилетевших особей с успешным гнездованием в этом местообитании в предыдущем году;
- индикаторы кормообеспеченности для специализированных видов-стенофагов (семеноядных и хищных птиц).
В горных условиях умеренных широт этот принцип маскируется низкой наглядностью ежегодной изменчивости разнокачественности территории по существенным для гнездования птиц параметрам и отсутствием ярко выраженных и понятных для исследователя релизеров. Но в то же время такие заметные ежегодные изменения условий обитания обнаруживаются в поймах крупных рек с периодическими изменениями паводкового режима, например, в пойме р. Оби (Ананин, 1986, 1989, 1995).
Сообщество птиц в горных условиях формируется и функционирует не по конкурентному, а по линдивидуалистическому типу (Пузаченко, Черенков, 2008), что определяется независимым друг от друга характером изменения плотности разных видов (межгодовых флуктуаций). Асинхронный характер колебаний обилия у разных видов способствует стабилизации плотности населения в целом.
Таким образом, население птиц того или иного местообитания можно рассматривать как результат индивидуального выбора всякой особью каждого вида подходящих для себя условий в конкретном году, поэтому плотность вида в локальном местообитании определяется следующими параметрами:
- метеорологические условия весны и начала лета (теплообеспеченность и количество осадков) и весенняя фенологическая обстановка (сроки схода снегового покрова и развития растительности), формирующие необходимые условия для гнездования;
- количество особей, достигших района гнездования и лосевших в конкретном местообитании, которое определяется во многом обстоятельствами пролонгации весенних миграций, сопровождающимися явлениями недолета или перелета, в первую очередь у видов, для которых район исследований расположен вблизи границы гнездового ареала, а также изменениями выживаемости на местах зимовок вследствие аномальных климатических и кормовых условий;
- для специализированных семеноядных и хищных видов птиц важна урожайность основных кормов или уровень численности видов-жертв в предыдущем и текущем году.
Выводы
1. Фауна птиц горных систем юга Восточной Сибири включает 378 видов, что составляет 93,3 % орнитофауны российской части бассейна оз. Байкал и 47,9 % авифауны России, в том числе гнездящиеся виды - 66,9 %, пролетные - 11,7 % и залетные - 20,6 %. Ее основу составляют, помимо широкораспространенных видов, представители сибирского, европейского, китайского и монгольского типов фаун, а по ареалогической структуре - виды палеарктической, голарктической и азиатской групп. Среди последних максимально представлены североазиатская и центральноазиатская подгруппы, а в меньшей степени - виды восточноазиатской и трансазиатской подгрупп. Авифаунистическое сходство между отдельными горными системами очень велико, что свидетельствует о единстве фауны птиц региона. Максимальные отличия выявлены между наиболее удаленными Баргузинским хребтом и Восточным Саяном.
2. Общий уровень плотности населения птиц в горах юга Восточной Сибири низкий. С увеличением высоты местности количество видов в населении выделов постепенно убывает, а обилие достигает своего максимума в предгорьях и подгольцово-субальпийском поясе. Напротив, наибольшее обилие зимующих птиц зарегистрировано в горно-лесном поясе, оно снижается в предгорных и равнинных лесах.
3. В горных условиях юга Восточной Сибири в последние десятилетия среди дальних мигрантов преобладают виды с отрицательным трендом долговременных изменений гнездового обилия, а у оседлых видов и ближних мигрантов соотношение количества видов с положительными и отрицательными трендами примерно равное. Максимальное сходство долговременных изменений обилия виды птиц имеют по местам преимущественных зимовок, географическое положение которых и складывающиеся в них условия влияют на обилие местного гнездового населения.
4. Метеорологические параметры весны и начала лета (теплообеспеченность и осадки), определяющие в горных условиях гнездопригодность и состояние кормовых ресурсов местообитаний, обуславливают формирование гнездового населения мигрирующих и оседлых видов птиц.
5. Сроки развития весенних фенологических процессов и параметры гидротермического режима весеннего сезона влияют на формирование местного населения птиц за счет изменения количества обосновавшихся в районе гнездования особей и вызывают разнонаправленные изменения обилия мигрирующих и оседлых видов в различных выделах с отличающимися условиями.
6. Изменение сроков прилета птиц существенно воздействует на обилие видов и может носить разнонаправленный характер в разных высотно-поясных выделах. Глобальные изменения климата, вследствие сдвига сроков весеннего прилета птиц, могут в горных условиях оказывать воздействие на формирование локального обилия гнездящихся видов.
7. Структура зимнего населения в горных условиях определяется в основном динамикой обилия немногих массовых видов (буроголовая гаичка, московка, обыкновенный поползень, белокрылый клест и обыкновенная чечетка).
8. Метеорологические показатели зимнего периода (соотношение морозности и снежности) не оказывают существенного воздействия на уровень плотности большинства зимующих видов птиц. Главный фактор, определяющий особенности пространственного распределения семеноядных птиц - урожайность семян основных кормовых растений. Для зимующих видов птиц не прослеживается тесная связь зимнего обилия с летней плотностью предыдущего года, что определяется высоким уровнем осенних кочевок. Летняя плотность некоторых оседлых видов в значительной мере обусловливается предшествующим уровнем их зимнего обилия.
9. Население птиц конкретного участка в горных условиях юга Восточной Сибири ежегодно формируется преимущественно из новых особей - иммигрантов. Этот процесс зависит от экологической обстановки, складывающейся в связи с условиями погоды весной и в начале лета, и соотношениями процессов эмиграции и иммиграции. К главным факторам, обуславливающим формирование локального видового обилия птиц, относятся метеорологические параметры (теплообеспеченность и осадки весной и в начале лета) и сроки наступления весенних фенологических явлений, а также общая численность особей, достигших района наблюдений.
Список основных публикаций автора по теме диссертации
Монографии
- Мониторинг природных комплексов Северо-Восточного Прибайкалья / А.А. Ананин, Т.Л. Ананина, Е.М. Черникин, Е.А. Дарижапов и др.: Тр. гос. прир. биосф. заповедника Баргузинский. - Вып. 8. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2002. - 224 с. (авторский вклад 35 %).
- Ананин А.А. Птицы Баргузинского заповедника / А.А. Ананин. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2006. - 276 с.
- Ананин А.А. Птицы Северного Прибайкалья: динамика и особенности формирования населения / А.А. Ананин. - Улан-Удэ: Изд-во Бурятского госуниверситета, 2010. - 296 с.
Статьи в рецензируемых журналах
- Ананин А.А. Значение весеннего паводка в динамике численности и разнообразия населения птиц средней Оби / А.А. Ананин // Сибирский экологический журнал. - 1995. - Т. 2. - № 2. - С. 137-145.
- Ананин А.А. Долговременные изменения сроков прилета птиц в Северо-Восточное Прибайкалье / А.А. Ананин // Сибирская орнитология. Вып. 4. / Вестник Бурятского университета. Спец. серия. - Улан-Удэ: Изд-во Бурятского гос. ун-та, 2006. - С. 7-17.
- Ананин А.А. Оценка долговременных тенденций и современное состояние совообразных Баргузинского хребта / А.А. Ананин // Вестник Бурятского гос. ун-та. Биология, география. - Улан-Удэ, 2008. - Вып. 4. - С. 139-144.
- Ананин А.А. Многолетняя динамика летнего населения птиц высокогорий Баргузинского хребта (Северо-Восточное Прибайкалье) /А.А. Ананин // Известия Самарского научного центра РАН. - 2009. - Т. 11. - № 1/1. - С. 9-12.
- Ананин А.А. Формирование видового населения птиц на высотном профиле западного макросклона Баргузинского хребта / А.А. Ананин // Вестник Бурятского гос. ун-та. Биология, география. - 2009. - Вып. 4. - С. 130-137.
- Ananin A.A. Long-term arrival trends of 54 avian species to Barguzinsky Nature Reserve in the northeastern Baikal area / A.A. Ananin, L.V. Sokolov // Avian. Ecol. Behav. - 2009. - V. 15. - P. 33-48. (авторский вклад 50 %).
- Ananin A.A. Relationship between weather conditions, crops of Siberian pine nuts, and irruptions of Siberian Nutcrackers Nucifraga caryocatactes macrorhynchos L. in Siberia and Europe / A.A. Ananin, L.V. Sokolov // Avian. Ecol. Behav. - 2009. - V. 15. - P. 23-31. (авторский вклад 50 %).
- Ананин А.А. Влияние абиотических факторов на динамику обилия зимующих видов птиц Баргузинского хребта / А.А. Ананин // Известия Иркутского гос. ун-та. Серия Биология. Экология. - 2010. - Т. 3. - № 4. - С. 45-51.
- Ананин А.А. Долговременная динамика чужеродных видов птиц Баргузинского заповедника / А.А. Ананин // Вестник Бурятской гос. сел.-хоз. академии. - № 3 (20). - 2010. - С. 70-75.
- Ананин А.А. Особенности формирования видового населения птиц западного макросклона Баргузинского хребта (Северо-Восточное Прибайкалье) / А.А. Ананин // Известия Самарского научного центра РАН. - 2010. - Т. 12 (33). - № 1 (5). - С. 1256-1259.
- Ананин А.А. Влияние фенологических сроков весны на формирование гнездового населения птиц в ландшафтно-зональных условиях гор Северо-Восточного Прибайкалья / А.А. Ананин // Вестник Томского государственного университета. Биология. - 2011. - № 4(16). - С. 66-79.
- Ананин А.А. Многолетняя динамика обилия фоновых видов птиц Баргузинского хребта в гнездовой период / А.А. Ананин // Вестник Бурятского гос. ун-та. Биология, география. - 2011. - Вып. 4. - С. 93-99.
- Ананин А.А. Многолетняя динамика плотности населения птиц и жужелиц на катене Баргузинского хребта (Северное Прибайкалье) / А.А. Ананин, Т.Л. Ананина // Известия Самарского научного центра РАН. - 2011. - Т. 13 (39). - № 1 (5). - С. 1041-1044. (авторский вклад 50 %).
Научные статьи в других изданиях и материалы конференций
- Ананин А.А. Влияние весеннего паводка на гнездящихся птиц в пойме Средней Оби / А.А. Ананин, Т.Л. Ананина // XVIII Международный Орнитологический Конгресс / Тез. докл. и стенд. сообщ. - М.: Наука, 1982. - С. 121-122. (авторский вклад 50 %).
- Ананин А.А. Редкие птицы Баргузинского заповедника / А.А. Ананин // Редкие, исчезающие и малоизученные птицы СССР / Сб. научн. тр. ЦНИЛ Главохоты РСФСР. - М., 1986. - С. 98-103.
- Ананин А.А. Пойменные циклы и кадастровые учеты птиц / А.А. Ананин // Всесоюзное совещание по проблеме кадастра и учета животного мира / Тез. докл. - Часть 2. - М., 1986. - С. 218-219.
- Ананин А.А. Роль весеннего паводка в динамике численности и структуры населения птиц Средней Оби / А.А. Ананин // Изучение птиц СССР, их охрана и рациональное использование / Тез. докл. IX Всесоюзн. орнитол. конф., 16-20 декабря 1986 г. - Часть 1. - Л., 1986. - С. 36-37.
- Ананин А.А. Численность и распределение зимующих птиц Северо-Восточного Прибайкалья / А.А. Ананин // Териология, орнитология и охрана природы / Тез. докл. XI Всесоюзн. симп. Биолог. проблемы Севера. Вып. 3. - Якутск, 1986. - С. 87-88.
- Равкин Ю.С. Пойменные циклы и кадастровые учеты птиц / Ю.С. Равкин, Ю.Ю. Лесневский, А.М. Адам, А.А. Ананин // Всесоюзное совещание по проблеме кадастра и учета животного мира / Тез. докл. - Часть 2. - М., 1986. - С. 218-219. (авторский вклад 25 %).
- Ананин А.А. Птицы / А.А. Ананин, А.В. Федоров // Фауна Баргузинского заповедника / Флора и фауна заповедников СССР. - М., 1988. - С. 8-33. (авторский вклад 50 %).
- Ананин А.А. Динамика численности птиц в изменчивых и стабильных местообитаниях и кадастровые учеты / А.А. Ананин // Всесоюзн. совещ. по проблеме кадастра и учета животного мира / Тез. докл. - Часть 3. - Уфа: Башкир. кн. изд-во, 1989. - С. 3-4.
- Ананин А.А. Изменение фауны птиц Баргузинского заповедника за счет расселяющихся видов / А.А. Ананин // Заповедники СССР - их настоящее и будущее / Тез. докл. Всесоюзн. конф. - Часть 3. Зоологические исследования. - Новгород, 1990. - С. 188-189.
- Ананин А.А. Динамика численности гнездового населения птиц Северо-Восточного Прибайкалья / А.А. Ананин // Материалы 10-й Всесоюзн. орнитол. конф., г. Витебск, 17-22 сентября 1991 г. / Тез. докл. - Часть 2. - Кн. 1. - Минск: Навука i тэхнiка, 1991. - С. 20-21.
- Ананин А.А. Изменения гнездовой численности птиц лесного пояса Баргузинского хребта / А.А. Ананин // Орнитологические проблемы Сибири / Тез. докл. к конф. - Барнаул, 1991. - С.115-116.
- Ананин А.А. Новые виды птиц Баргузинского заповедника / А.А. Ананин // Русский орнитол. журн. - 1995. - Т. 4. - Вып. 1/2. - С. 64-65.
- Ананин А.А. Организация орнитологического мониторинга в Баргузинском заповеднике / А.А. Ананин // Состояние и проблемы особо охраняемых природных территорий Байкальского региона / Матер. республ. совещания. - Улан-Удэ: Бурят. гос. ун-т, 1996. - С. 5-7.
- Ананин А.А. Орнитологический мониторинг в Баргузинском заповеднике / А.А. Ананин // Биологическое разнообразие животных Сибири: Матер. науч. конф. - Томск, 1998. - С. 178-179.
- Ананин А.А. Результаты долговременного изучения фауны и населения птиц Баргузинского заповедника / А.А. Ананин // Беловежская пуща на рубеже третьего тысячелетия : Матер. науч.-практ. конф. - Минск: БГУ, 1999. - С. 248-250.
- Ананин А.А. Долговременные изменения сроков весеннего и осеннего пролета птиц на территории Баргузинского заповедника / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии : Матер. I Междунар. орнитол. конф. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2000. - С.103-104.
- Ананин А.А. Многолетняя динамика численности зимнего населения птиц Баргузинского заповедника / А.А. Ананин // Анализ многолетних рядов наблюдений за природными компонентами в заповедниках Дальнего Востока. - Владивосток: Дальнаука, 2000. - С.4-18.
- Ананин А.А. Многолетняя динамика численности летнего населения птиц Баргузинского заповедника / А.А. Ананин // Экология и рациональное природопользование на рубеже веков. Итоги и перспективы : Матер. междунар. науч. конф. - Том II. - Томск, 2000. - С.8-10.
- Ананин А.А. Орнитологический мониторинг в Баргузинском заповеднике / А.А. Ананин // Инвентаризация, мониторинг и охрана ключевых орнитологических территорий России / Матер. совещаний по программе "Ключевые орнитологические территории России" (1998-2000 гг.). - Вып. 2. - М., 2000. - С.65-76.
- Ананин А.А. Орнитофаунистические исследования в Северо-Восточном Прибайкалье / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии : Матер. I Междунар. орнитол. конф. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2000. - С.40-42.
- Ананин А.А. Итоги и перспективы орнитологического мониторинга в Баргузинском биосферном заповеднике / А.А. Ананин // Современные проблемы биоиндикации и биомониторинга / Тез. докл. XI Междунар. симпоз. по биоиндикаторам. - Сыктывкар, 2001. - С. 6 (рус.). - С. 220-221 (англ.).
- Ананин А.А. Многолетняя динамика численности летнего и зимнего населения птиц Баргузинского заповедника / А.А. Ананин // Достижения и проблемы орнитологии Северной Евразии на рубеже веков / Тр. Междунар. конф. Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Евразии. - Казань: Изд-во Магариф, 2001. - С. 295-316.
- Ананин А.А. Подготовка баз данных долговременного мониторинга состояния природных комплексов Баргузинского заповедника / А.А. Ананин // ГИС в научных исследованиях заповедников Сибири : Тр. междунар. науч. конф., посвящ. 75-летию гос. заповедника Столбы. - Красноярск, 2001. - С. 5-16.
- Ананин А.А. Общий обзор фауны птиц Северо-Восточного Прибайкалья (Баргузинский хребет) / А.А. Ананин // Тр. гос. природ. заповедника Байкало-Ленский. - Вып. 2. - Иркутск, Листок, 2001. - С. 66-82.
- Ананин А.А. Влияние изменений климата на фенологию птиц в Баргузинском заповеднике / А.А. Ананин // Многолетняя динамика численности птиц и млекопитающих в связи с глобальными изменениями климата : Матер. междунар. симпозиума (11-16 ноября 2002, Россия, Республика Татарстан, Казань). - Казань: ЗАО Новое знание, 2002. - С.107-112.
- Ананин А.А. Долговременные изменения сроков весеннего прилета птиц на территории заповедника Баргузинский / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии: Матер. II Междунар. орнитол. конф. - Ч. 1. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2003. - С. 143-147.
- Ананин А.А. Редкие виды птиц Баргузинского заповедника / А.А. Ананин // Структура и функционирование экосистем Байкальской Сибири : Матер. регион. науч.-практ. конф. (24 октября 2002 г.). - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2003. - С.65-67.
- Ананин А.А. Долговременная динамика летней и зимней численности птиц на западном макросклоне Баргузинского хребта / А.А. Ананин // Сибирская зоологическая конференция / Тез. докл. Всерос. конф., посвященной 60-летию Института систематики и экологии животных СО РАН, 15-22 сентября 2004 г. - Новосибирск, 2004. - С. 99-100.
- Ананин А.А. Роль заповедников в орнитологическом мониторинге климатических изменений / А.А. Ананин // Актуальные проблемы управления заповедниками в Европейской части России : Матер. юбилейной науч.-практич. конф. - Воронеж: Воронеж. гос. ун-т, 2004. - С. 58-61.
- Фефелов И.В. Динамика численности мигрирующих уток в Байкальских дельтах в 1976-1996 гг. / И.В. Фефелов, В.С. Садков, А.А. Ананин, И.И. Тупицын, В.А. Подковыров, А.В. Шинкаренко, Н.Н. Сафронов, Н.Г. Скрябин, В.Е. Журавлев // Казарка № 10 / Бюлл. рабоч. группы по гусеобразным Северной Евразии. - М., 2004. - С. 323-338. (авторский вклад 15 %).
- Ананин А.А. Динамика населения серых журавлей в Баргузинском заповеднике / А.А. Ананин // Результаты охраны и изучения природных комплексов Сихотэ-Алиня / Матер. междунар. науч.-практич. конф. - Владивосток: ОАО Примполиграфкомбинат, 2005. - С. 355-360.
- Ананин А.А. Мониторинг дневных хищных птиц в Баргузинском заповеднике / А.А. Ананин // Всерос. конф. Природная и антропогенная динамика наземных экосистем, посвященная памяти выдающегося исследователя лесов Сибири А.С. Рожкова (1925-2005 гг.): Матер. Всерос. конф. (Иркутск, 11-15 октября 2005 г.). - Иркутск: Изд-во Иркут. гос. техн. ун-та, 2005. - С. 154-157.
- Ананин А.А. Мониторинг куриных птиц в Баргузинском заповеднике / А.А. Ананин // Охрана и рациональное использование животных и растительных ресурсов : Матер. междунар. науч.-практ. конф., Иркутск, 26-30 мая 2005 г. - Иркутск: Изд-во ИрГСХА, 2005. - С. 198-202.
- Ананин А.А. Влияние погодно-климатических изменений на сроки прилета и гнездовую плотность массовых видов птиц Баргузинского заповедника (Северо-Восточное Прибайкалье) / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии. - Вып. 3. - Ч. 1. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2006. - С. 6-8.
- Ананин А.А. Динамика встречаемости хищных птиц и сов на территории Баргузинского заповедника в 1984-2004 гг. / А.А. Ананин // Региональные проблемы заповедного дела : Матер. междунар. науч.-практ. конф. - Абакан: Изд-во Хакасского гос. ун-та, 2006. - С. 213-215.
- Ананин А.А. Динамика численности водоплавающих птиц Баргузинского заповедника / А.А. Ананин // Природные комплексы Баргузинского хребта : Тр. гос. природ. биосферн. заповедника Баргузинский. - Вып. 9. - Улан-Удэ: Изд-во БГУ, 2006. - С. 44-63.
- Ананин А.А. Долговременные исследования динамики численности птиц Баргузинского хребта / А.А. Ананин // Развитие современной орнитологии в Северной Евразии: Тр. XII Междунар. онитол. конф. Северной Евразии. - Ставрополь: Изд-во СГУ, 2006. - С. 280-297.
- Ананин А.А. Долговременный мониторинг гнездовой популяции вальдшнепа (Scolopax rusticola) в Баргузинском заповеднике / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии. - Вып. 3. - Ч. 2. - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2006. - С. 26-29.
- Ананин А.А. Особенности долговременной динамики численности кукши (Perisoreus infaustus) в Северо-Восточном Прибайкалье / А.А. Ананин // Популяционная экология животных : Матер. Междунар. конф. Проблемы популяционной экологии животных, посвящ. памяти акад. И.А. Шилова. - Томск: Томский гос. ун-т, 2006. - С. 51.
- Беляев К.Г. Результаты изучения миграций воробьиных птиц на севере озера Байкал / К.Г. Беляев, А.А. Ананин // Природные комплексы Баргузинского хребта: Тр. гос. природ. биосферн. заповедника Баргузинский. - Вып. 9. - Улан-Удэ: Изд-во БГУ, 2006. - С. 63-138. (авторский вклад 50 %).
- Ананин А.А. Долговременный мониторинг летнего населения птиц высокогорий Баргузинского хребта (Северо-Восточное Прибайкалье) / А.А. Ананин // Биоразнообразие и сохранение генофонда флоры, фауны и народонаселения Центрально-Азиатского региона : матер. 2-ой междунар. науч.-практич. конф. 26-29 сент. 2007 г. - Кызыл, 2007. - С. 115-117.
- Ананин А.А. Долговременная динамика численности летнего населения птиц высокогорий Баргузинского хребта (Северо-Восточное Прибайкалье) / А.А. Ананин // Горные экосистемы и их компоненты : Тр. междунар. конф. - Ч. 1. - М.: Т-во научных изданий КМК, 2007. - С. 47-52.
- Ананин А.А. Многолетний мониторинг летнего населения птиц высокогорий Баргузинского хребта (Северо-Восточное Прибайкалье) / А.А. Ананин // Биоразнообразие, проблемы экологии Горного Алтая и сопредельных регионов : настоящее, прошлое, будущее / Матер. Междунар. конф. - Часть 1. - Горно-Алтайск: РИО ГАГУ, 2008. - С. 5-10.
- Ананин А.А. Результаты долговременного мониторинга хищных птиц в Северо-Восточном Прибайкалье / А.А. Ананин // Изучение и охрана хищных птиц Северной Евразии : Матер. V Междунар. конф. по хищным птицам Северной Евразии. - Иваново, 2008. - С.172-174.
- Ананин А.А. Формирование населения птиц в горных условиях / А.А. Ананин // Биология : теория, практика, эксперимент : матер. Междунар. науч. конф. - Саранск, 2008. - Кн. 1. - С. 82-84.
- Ананин А.А. Влияние фенологических сроков весны на изменения численности птиц в Северо-Восточном Прибайкалье / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии : матер. IV Междунар. орнитол. конф. (17-20 сентября 2009 г.) - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2009. - С. 120-124.
- Ананин А.А. Пространственно-временная изменчивость населения птиц западного макросклона Баргузинского хребта (Северо-Восточное Прибайкалье) / А.А. Ананин // Животный мир горных территорий. - М.: Т-во научных изданий КМК, 2009. - С. 181-184.
- Ананин А.А. Связь динамики численности видового населения птиц со сроками прилета в район гнездования / А.А. Ананин // Современные проблемы орнитологии Сибири и Центральной Азии : матер. IV Междунар. орнитол. конф. (17-20 сентября 2009 г.) - Улан-Удэ: Изд-во Бурят. гос. ун-та, 2009. - С. 16-19.
- Ананин А.А. Долговременная изменчивость населения птиц западного макросклона Баргузинского хребта / А.А. Ананин // Орнитогеография Палеарктики: современные проблемы и перспективы / Под ред. Ю.С. Равкина, Г.С. Джамирзоева и С.А. Букреева. - Махачкала, 2009. - С. 102-109.
- Ананин А.А. Долговременные изменения фауны птиц Баргузинского заповедника / А.А. Ананин // Биодиверситиология : Современные проблемы сохранения и изучения биологического разнообразия / Сб. матер. II Междунар. науч.-практ. конф. - Чебоксары: типография Новое время, 2010. - С. 64-67.
- Ананин А.А. Особенности долговременной динамики населения пеночек на Баргузинском хребте / А.А. Ананин // Биоразнообразие, проблемы экологии Горного Алтая и сопредельных регионов: настоящее, прошлое, будущее / Матер. II Междунар. конф. 20-24 сентября 2010 г. - Горно-Алтайск: РИО ГАГУ, 2010. - С. 5-9.
- Ананин А.А. Особенности долговременной динамики населения пеночки-зарнички на западном макросклоне Баргузинского хребта / А.А. Ананин // Актуальные вопросы изучения птиц Сибири: матер. Сибирской орнит. конф., посвящ. памяти и 75-летию Э.А. Ирисова. - Барнаул: Азбука, 2010. - С. 54-57.
- Ананин А.А. Особенности зимнего и летнего распределения зимующих видов птиц Баргузинского хребта / А.А. Ананин // Горные экосистемы Южной Сибири: изучение, охрана и рациональное природопользование / Тр. Тигирекского заповедника. - Вып. 3. - Барнаул, 2010. - С. 262-264.
- Ананин А.А. Особенности формирования видового населения птиц на высотном профиле западного макросклона Баргузинского хребта / А.А. Ананин // Орнитология в Северной Евразии. Матер. XIII Междунар. орнитол. конф. Северной Евразии. Тез. докл. / Отв. ред. Е.Н. Курочкин, А.В. Давыгора. - Оренбург: изд-во Оренбургского гос. пед. ун-та, ИПК ГОУ ОГУ, 2010. - С. 32.
- Ананин А.А. Птицы как объект долговременного мониторинга природных комплексов ООПТ / А.А. Ананин // Антропогенная трансформация природной среды : матер. междунар. конф. (18-21 октября 2010 г.) / Перм. гос. ун-т. - Пермь, 2010. - Т. 2. - С. 8-14.
- Ананин А.А. Влияние долговременных климатических изменений на формирование населения птиц Баргузинского хребта / А.А. Ананин // Разнообразие почв и биоты Северной и Центральной Азии : матер. II междунар. науч. конф. - Т. 3. - Улан-Удэ: Изд-во БН - СО РАН, 2011. - С. 97-98.
- Ананин А.А. Долговременная динамика численности и сроков миграций куликов на северо-восточном побережье озера Байкал / А.А. Ананин // Кулики Северной Евразии : экология, миграции и охрана: Матер. VIII Междунар. науч. конф. (10-12 ноября 2009 г., Ростов-на-Дону). - Ростов н/Д: Изд-во ЮН - РАН, 2011. - С. 217-240.
- Ананин А.А. Долговременные изменения гнездового обилия фоновых видов птиц Баргузинского хребта / А.А. Ананин // Современные проблемы биологии и экологии : матер. докл. Междунар. науч.-практич. конф., 10-12 марта 2011 г. - Махачкала: ДГПУ, 2011. - С. 119-122.
- Ананин А.А. Долговременный мониторинг куриных птиц в Баргузинском заповеднике // Сохранение разнообразия животных и охотничье хозяйство России: Матер. 4-й Междунар. науч.-практич. конф. - М.: Изд-во РГАУ-МСХА им. К.А. Тимирязева, 2011. - С. 349-351.
Все авторефераты докторских диссертаций