
 Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]
Защитное действие модуляторов эндогенной каннабиноидной системы при экспериментальной церебральной ишемии
Автореферат кандидатской диссертации
На правах рукописи
ГЕНРИХС ЕЛИЗАВЕТА ЕВГЕНЬЕВНА
ЗАЩИТНОЕ ДЕЙСТВИЕ МОДУЛЯТОРОВ ЭНДОГЕННОЙ КАННАБИНОИДНОЙ СИСТЕМЫ
ПРИ ЭКСПЕРИМЕНТАЛЬНОЙ ЦЕРЕБРАЛЬНОЙ ИШЕМИИ
03.03.04 - Клеточная биология, цитология, гистология
а
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва - 2012
Работа выполнена в Федеральном Государственном бюджетном учреждении Научный центр неврологии Российской академии медицинских наук
Научный руководитель:
доктор биологических наук Хаспеков Леонид Георгиевич
Научный консультант:
кандидат химических наука Бобров Михаил Юрьевич
Официальные оппоненты:
доктор биологических наук, профессор, аа Мустафин Александр Газисович
профессор кафедры биологии педиатрического
факультета ГБОУ ВПО Российского национального
исследовательского медицинского университета им.
Н.И. Пирогова Минздравсоцразвития России
доктор биологических наук, а ааПлотников Егор Юрьевич
ведущий научный сотрудник НИИ физико-
химической биологии им. А.Н. Белозерского
Московского Государственного
университета им. М.В. Ломоносова
Ведущая организация:а аФГБУ Научно-исследовательский институт морфологии человека РАМН
Защита состоится л28 мая 2012 года в 12:00 на заседании диссертационного совета
Д 208.072.04 на базе Российского национального исследовательского медицинского университета им. Н.И. Пирогова Минздравсоцразвития РФ адресу: 117997, г. Москва, ул. Островитянова, д. 1.
С диссертацией можно ознакомиться в библиотеке Российского национального исследовательского медицинского университета им. Н.И. Пирогова Минздравсоцразвития РФ по адресу: 117997, Москва, ул. Островитянова, д. 1
Автореферат разослан л25 апреля 2012 года
Ученый секретарь диссертационного совета
доктор медицинских наук, профессор а .В. Леонова
![]()
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования.
Острые нарушения мозгового кровообращения, как и многие другие заболевания центральной нервной системы, становятся в последние годы все более актуальной медико-социальной проблемой. Инсульт занимает одно из ведущих мест среди причин смертности и всегда первое место среди причин стойкой утраты трудоспособности. Изучение динамики заболеваемости за последние десятилетия указывает на существование устойчивой тенденции к росту (0,5-1 % в год) числа острых нарушений мозгового кровообращения.
Высокая медицинская и социальная значимость острой сосудистой патологии головного мозга определяет приоритетность научных исследований, связанных с созданием оптимальных схем лечения, а также возможностью подбора препаратов, осуществляющих нейрозащитное действие в условиях ишемии. Существующая неудовлетворенность результатами современной комплексной терапии ишемии мозга объясняется сохранением высокого уровня летальности, значительной инвалидизацией и социальной дезадаптацией пациентов, перенесших инсульт, что во многом связано с отсутствием в арсенале клиницистов эффективных нейропротекторов (Верещагин с соавт., 2002). Данное обстоятельство заставляет более детально изучать механизмы ишемического повреждения структур мозга, а также стимулирует на поиск новых фармакологических агентов для оптимизации всего комплекса лечения больных с инсультами с целью более продуктивного оказания экстренной помощи, снижения летальности, сокращения длительности острого периода, ранней активизации больных и уменьшения инвалидизирующих последствий заболевания.
В качестве перспективной группы соединений для разработки потенциальных нейропротекторов нами были выбраны эндоканнабиноиды (ЭК) и их синтетические аналоги. Эндоканнабиноиды и их рецепторы, входящие в состав эндогенной каннабиноидной системы (ЭКС), обеспечивают тонкую регуляцию синаптической активности и пластичности за счет снижения выброса большинства известных нейромедиаторов, в том числе возбуждающего нейромедиатора - глутамата (Kogan and Mechoulam, 2007). Поэтому представляется целесообразной разработка на основе модуляторов эндогенной каннабиноидной системы новых терапевтических подходов к коррекции повреждений нервной ткани, связанных с нарушениями регуляции нейромедиаторных процессов.
Ведущую роль в запуске каскада цитотоксических реакций, приводящих к ишемической гибели клеток при острых расстройствах мозгового кровообращения, играет чрезмерная стимуляция рецепторов глутамата, которая вызывается его усиленным выбросом из пресинаптических терминалей и приводит к накоплению в нейронах ионов кальция, поступающих через ионные каналы. Стойкое повышение внутриклеточной концентрации Са2+ инициирует деструктивные липо- и протеолитические реакции, сопровождается генерацией свободных радикалов и реактивных форм кислорода, что в конечном итоге приводит к необратимым повреждениям нервных клеток по пути некроза и/или апоптоза. Повреждению нервной ткани сопутствуют воспалительные реакции, которые также усугубляют течение нейродеструктивного процесса (Гусев с соавт., 1997).
Одним из наиболее изученных эндоканнабиноидов является N-арахидоноилэтаноламин (анандамид). Было показано увеличение содержание анандамида и его предшественника в ткани мозга при ишемическом воздействии (Panikashvili et al., 2001; Schabitz et al., 2002). Анандамид защищает нейроны в культуре от цитотоксического действия глутамата (Shen and Thayer, 1998), а также снижает выраженность ишемических повреждений в некоторых моделях ишемии in vivo (Schabitz et al., 2002; Franklin et al., 2003; Muthian et al., 2004; Degn et al., 2007; Amantea et al., 2007), что свидетельствует о несомненном нейропротекторном потенциале представителей семейства эндоканнабиноидов.
Важно отметить, что в тканях млекопитающих были обнаружены и другие производные жирных кислот, взаимодействующие с белками эндоканнабиноидной системы. На сегодняшний день это довольно большая группа соединений, представленная преимущественно амидами ненасыщенных жирных кислот с биогенными аминами (этаноламин, аминокислоты, нейротрансмиттеры). Физиологическая роль подавляющего большинства представителей данного семейства и их метаболитов остается до конца не изученной.
Недавно из мозга крысы был выделен новый эндоканнабиноид N-арахидоноилдофамин (АД). В ряде работ было показано, что АД и другие N-ацилдофамины способны ингибировать агрегацию тромбоцитов, стимулируют мозговую микроциркуляцию (Васильева с соавт., 2002), проявляют противовоспалительное и антиоксидантное действие (Бобров с соавт., 2006), однако нейрозащитное действие N-ацилдофаминов практически не изучалось.
Существенным недостатком анандамида и родственных соединений является их низкая метаболическая стабильность и, как следствие, быстрая инактивация. Под действием различных клеточных гидролаз, в частности гидролазы амидов жирных кислот (ГАЖК), анандамид распадается на арахидоновую кислоту и этаноламин (Vandevoorde and Lambert, 2007). N-ацилдофамины также являются субстратом этого фермента, хотя их гидролиз протекает с меньшей скоростью. Поэтому использование ингибиторов гидролазы амидов жирных кислот может рассматриваться как способ повышения эффективности действия эндоканнабиноидов с нейропротекторными свойствами.
Таким образом, изучение механизмов действия агонистов каннабиноидных рецепторов позволит расширить представление о функционировании и роли ЭКС в организме, а также разработать на основе модуляторов ЭКС новые терапевтические подходы к коррекции повреждений нервной ткани, связанных с нарушениями регуляции нейромедиаторных процессов.
Цель работы.
Исследование нейропротекторных свойств эндоканнабиноидов семейства N-ацилдофаминов, а также комбинации N-ацилдофаминов с ингибиторами их гидролиза на моделях повреждений нейронов в культурах клеток различных структур головного мозга и при экспериментальной церебральной ишемии in vivo.
Задачи исследования.
- Оценить на светооптическом уровне влияние N-ацилдофаминов N-арахидоноилдофамина и N-докозагексаеноилдофамина на степень повреждения нейронов при индукции апоптоза и цитотоксическом действии глутамата в культурах клеток мозжечка и новой коры, исследовать нейропротекторный эффект этих соединений, а также выявить возможные механизмы этого эффекта.
- Исследовать развитие культуры клеток гиппокампа на мультиэлектродной матрице и оценить влияние каннабиноидов N-арахидоноилдофамина и N-докозагексаеноилдофамина на вызванные глутаматом изменения биоэлектрической активности нейронов в функциональной нейронной сети, сформированной клетками гиппокампа.
- Изучить влияние на выживаемость клеток N-ацилдофаминов, используемых совместно с ингибиторами их гидролиза в модели нейронального апоптоза in vitro.
- Провести патоморфологическое исследование зоны ишемического повреждения коры головного мозга крыс при действии модуляторов ЭКС и выявить их влияние на ультраструктурные изменения в клетках при фокальной ишемии, индуцированной фототромбозом.
Научная новизна.
В результате проведенного исследования получены новые данные о нейропротекторных свойствах и возможных механизмах защитного действия модуляторов эндоканнабиноидной системы из семейства N-ацилдофаминов при моделировании апоптотического повреждения и цитотоксическом действии глутамата в культуре клеток различных структур головного мозга. Применение современной технологии культивирования клеток на мультиэлектродной матрице позволило показать роль активации каннабиноидных рецепторов (КР) в нормализации биоэлектрической активности нейронной сети, подвергнутой цитотоксическому воздействию глутамата. Впервые получены данные о нейрозащитном действии N-ацилдофаминов при ишемическом повреждении коры головного мозга крыс. Предложен подход для повышения эффективности нейрозащитного действия за счет применения N-ацилдофаминов в комбинации с ингибиторами их гидролиза.
Научно-практическая значимость.
Полученные данные расширяют представления о механизмах нейропротекторного действия модуляторов ЭКС и могут служить экспериментальным обоснованием разработки фармакологических агентов, способных уменьшать или предотвращать нейродегенеративные изменения при ишемических формах церебральной патологии, а также активировать репаративные процессы в постишемический период.
Положения, выносимые на защиту.
1. Исследованные N-ацилдофамины при индукции апоптоза и цитотоксическом действии глутамата в культуре клеток мозжечка и новой коры оказывают дозозависимый нейропротекторный эффект, опосредуемый каннабиноидными рецепторами 2-го типа.
2.а N-арахидоноилдофамин и N-докозагексаеноилдофамин нормализуют вызванные глутаматом изменения биоэлектрической активности нейронов в функциональной нейронной сети, сформированной клетками гиппокампа на мультиэлектродной матрице.
3. Повышение нейропротекторного потенциала модуляторов эндогенной каннабиноидной системы достигается благодаря совместному использованию агонистов каннабиноидных рецепторов и ингибиторов их гидролиза.
4. Исследованные модуляторы эндогенной каннабиноидной системы оказывают нейропротекторный эффект и активируют репаративные процессы в коре головного мозга крыс при ее ишемическом повреждении in vivo.
Внедрение результатов исследования.
Перспективой внедрения результатов исследования является возможность их использования в материалах семинаров и лекций для студентов биологических и медицинских специальностей и школ для молодых ученых по вопросам экспериментальной и клинической неврологии. Исследованные соединения могут послужить прототипами новых лекарственных средств коррекции нейродеструктивных процессов при ишемических формах церебральной патологии.
Апробация диссертации.
Работа апробирована и рекомендована к защите на совместном заседании лабораторий Отдела исследований мозга Научного центра неврологии РАМН, НИИ Физико-химической биологии им. А.Н. Белозерского МГУ, Института биоорганической химии им. акад. М.М. Шемякина и Ю.А. Овчинникова РАН (ноябрь 2011 г.). Материалы диссертации были представлены на XIII научной школе-конференции молодых ученых по физиологии высшей нервной деятельности и нейрофизиологии (Москва, 2009), IV Международной научной конференции молодых ученых-медиков (Курск, 2010), 6-м Международном междисциплинарном конгрессе Нейронаука для медицины и психологии (Судак, Украина, 2010), Всероссийской конференции Современные направления в исследовании функциональной межполушарной асимметрии и пластичности мозга (Москва, 2010), конференции молодых ученых НЦН РАМН (Москва, 2011), Всероссийской конференции Рецепторы и внутриклеточная сигнализация (Пущино, 2011), 3-м Международном симпозиуме "Topical Problems of Biophotonics-2011" (Нижний Новгород, 2011).
Публикации.
По теме диссертации опубликовано 9 печатных работ, в том числе 2 статьи в рецензируемых журналах, рекомендованных ВАК РФ, 5 - в сборниках статей, 2 тезиса докладов на международных конференциях.
Объём и структура работы.
Диссертация изложена на 144 страницах машинописного текста, содержит 5 таблиц и иллюстрирована 24 фотографиями и рисунками. Работа включает введение, обзор литературных данных, материалы и методы исследования, результаты собственных исследований, их обсуждение, выводы и библиографический указатель, состоящий из 19 отечественных и 246 зарубежных источников.
_________________________________________
Автор выражает искреннюю благодарность сотрудникам Отдела исследований мозга ФГБУ НЦН РАМН и научной проблемной группы Клеточные технологии НИИ прикладной и фундаментальной медицины Нижегородской Государственной медицинской академии под руководством д.б.н., профессора И.В.Мухиной.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Основные правила содержания и ухода за экспериментальными животными соответствовали нормативам, данным в Приказе Минздрава России № 267 от 19.06.03 Об утверждении правил лабораторной практики в Российской Федерации. Исследование было одобрено этическим комитетом. Эндоканнабиноиды и их аналоги были синтезированы и охарактеризованы в лаборатории оксилипинов ИБХ РАН.
В экспериментах in vitro использованы: первичные монослойные культуры клеток мозжечка 7-8 дневных крыс линии Wistar, первичные монослойные культуры клеток новой коры и гиппокампа 16-18 дневных эмбрионов мышей линии С57Black; органотипические эксплантаты гиппокампа 7-9 дневных мышей линии С57Black. В исследовании было использовано свыше 250 культур.
Клетки мозжечка, новой коры и гиппокампа получали ферментно-механической диссоциацией соответствующих тканей по опубликованной ранее методике (Викторов, 1987). Суспензию клеток мозжечка и коры в питательной среде помещали в лунки 24- или 48-луночных пластиковых планшетов, покрытых опорным субстратом полиэтиленимином. Плотность клеток составляла 3-5 х103 клеток на мм2. Культуры инкубировали в СО2-инкубаторе (5 % СО2, 95 % воздуха), при температуре 35,5? С и относительной влажности 98%. Культуры клеток мозжечка использовали в экспериментах через 7 дней культивирования, а клетки новой коры через 11-12 дней инкубации.
Клетки гиппокампа культивировали на мультиэлектродной матрице системы MED64 в СО2-инкубаторе 14 дней (Мухина с соавт., 2009; Мухина и Хаспеков, 2010). Плотность клеток составляла 1-3 х103 клеток на мм2. Биоэлектрическую активность нейронов оценивали с использованием системы регистрации импульсов мультиэлектродных матриц (MED64, Япония) на базе НИИ прикладной и фундаментальной медицины Нижегородской Государственной медицинской академии (г. Нижний Новгород).
Для получения органотипических эксплантатов гиппокампа использовали методику, опубликованную ранее (Stoppini et al., 1991). Тканевые срезы толщиной 300 мкм, приготовленные из гиппокампа 7-8 дневных мышей с помощью микротома (McIlwain Tissue Chopper, IntraCell), помещали на поликарбонатные полупроницаемые мембраны (Millipore), по 4-5 срезов на мембрану. Мембраны вносили в лунки 6-луночного планшета, содержащие питательную среду, и культивировали в СО2-инкубаторе в течение 12-14 дней.
Для иммуногистохимической идентификации каннабиноидных рецепторов 1-го и 2-го типа (КР1 и КР2) на клетках мозжечка культуры, фиксированные в 4 % формальдегиде, обрабатывали первичными антителами к КР1 или КР2 (козьи поликлональные антитела Santa Cruz, США) в течение ночи при 4? С. Затем культуры обрабатывали 30 мин вторичными антителами, конъюгированными с флуоресцеинизотиоцианатом (антитела осла IgG-FITC, Santa Cruz, США). Далее препараты заключали в глицерин и исследовали методом флуоресцентной микроскопии. Контрольные культуры первичными антителами не обрабатывали.
Защитные свойства эндоканнабиноидов и их аналогов исследовали на моделях нейронального апоптоза и цитотоксического действия глутамата. Апоптоз индуцировали снижением концентрации К+ в питательной среде с 25 мМ до 9 мМ (К+-депривация). Одновременно в инкубационные среды добавляли исследуемые вещества и инкубировали 24 часа. Цитотоксическое действие достигалось добавлением к культурам глутамата в концентрации 100 мкМ на 15 минут. Затем глутамат удаляли и клетки инкубировали в присутствии исследуемых модуляторов ЭКС в течение 16-20 часов. Жизнеспособность клеток в обеих моделях изучали с помощью световой фазовоконтрастной микроскопии и оценивали количественно методом спектрофотометрического биохимического анализа (МТТ-тест, Mosmann T., 1983).
Срезы гиппокампа обрабатывали глутаматом 200 мкМ в течение 15 минут, после чего глутамат удаляли, а в инкубационные среды добавляли исследуемые вещества на 24 ч. Выживаемость срезов оценивали с помощью флуоресцентного красителя пропидий иодида (PI), который использовали как индикатор повреждения клеток (Kristensen et al., 2001). Окраска погибших клеток визуализировалась с помощью инвертированного флуоресцентного микроскопа. Срезы фотографировались как до эксперимента, так и после цитотоксического воздействия. Каждый эксперимент повторялся 2-3 раза в 4 сериях.
К клеткам, культивированным на мультиэлектродной матрице, добавляли последовательно вещества (глутамат и агонисты каннабиноидных рецепторов в различных концентрациях), регистрируя внеклеточные потенциалы действия в виде спонтанной биоэлектрической активности или спайковой. Оценивали электрическую активность по количеству пачек импульсов и количеству спайков в пачке.
В экспериментах in vivo для исследования нейропротекторного эффекта исследуемых соединений, а также их комбинации (лиганда КР и ингибитора ГАЖК) использовалась методика моделирования фокальной ишемии головного мозга крыс, индуцированной фототромбозом (ФТ) кровеносных сосудов новой коры по опубликованной ранее методике (Watson et al., 1988) в модификации И.В. Викторова (Островская с соавт., 1998). Экспериментальным животным через 30 минут после индукции ФТ и затем ежесуточно однократно в течение 4-х суток вводили внутрибрюшинно N-докозагексаеноилдофамин (5 мг/кг) или его же (в концентрации 2,5 мг/кг) в сочетании с ингибитором ГАЖК N-эйкозапентаеноилсеротонином (5 мг/кг). Далее мозг фиксировали и готовили серийные срезы толщиной 100 мкм, которые окрашивали 0,2 % раствором метиленового синего для морфометрического анализа с использованием компьютерных программ анализа изображения. Для гистологического анализа срезы мозга окрашивали гематоксилином с докрашиванием ванадиевым кислым фуксином (Викторов, Барсков, 1994).
Для исследования ультраструктурных характеристик перифокальной зоны очага, индуцированного фототромбозом, ее фрагменты, непосредственно прилежащие к зоне некроза, обрабатывали по стандартной методике, используемой для электронной микроскопии нервной ткани (Боголепов, 1976), и исследовали в электронном микроскопе (Hitachi).
Статистическая обработка полученных результатов проводилась на персональном компьютере с использованием пакетов программ Microsoft Excel XP, ConductorЩ (Alpha MED Sciences, Япония), прикладных программ Matlab и MEAMAN, BioStat (AnalystSoft).
Для характеристики полученных данных использовались критерии описательной статистики: определение средних значений и стандартного отклонения. Достоверность различий оценивали с помощьюа однофакторного дисперсионного анализа, применялись критерии Вилкоксона и Вилкоксона-Манна-Уитни. Статистически значимыми считали результаты при р<0,05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
1. Нейропротекторное действие модуляторов ЭКС in vitro.
1.1. Иммуногистохимическая идентификация каннабиноидных рецепторов в культуре клеток мозжечка.
Адекватность использования клеточных культур в исследованиях роли ЭКС в защитных механизмах подтверждена нами на основе иммуногистохимической идентификации (двойное мечение) каннабиноидных рецепторов КР1 и КР2, экспрессируемых культивируемыми клетками мозжечка. Были использованы первичные антитела к КР1 и КР2 и вторичные антитела, конъюгированные с флуорохромом FITС (Santa Cruz, США).
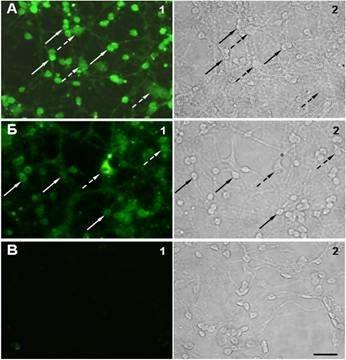
В случае обработки культур антителами к КР1 или КР2 интенсивный сигнал наблюдался как на нейронах, так и на глиальных клетках (рис. 1), однако при окраске антителами к КР1 наиболее интенсивный сигнал был на нейронах (рис. 1A), а к КР2 - преимущественно на глиальных клетках, среди которых присутствовали клетки с довольно интенсивным окрашиванием (рис. 1Б). Полученные данные позволяют предположить, что КР1 экспрессируются преимущественно нейронами, тогда как КР2 - глиальными клетками.
|
Рис. 1. Иммуногистохимическая визуализация каннабиноидных рецепторов 1-го типа (А1) и каннабиноидных рецепторов 2-го типа (Б1) в культуре клеток мозжечка. В1 - контрольная культура, окрашенная только вторичными антителами. А2-В2 - соответствующие поля зрения в фазовом контрасте. Сплошными стрелками показаны нейроны, пунктирными - глиальные клетки. Масштаб 20 мкм. 7 дней in vitro. |
1.2. Нейропротекторное действие N-ацилдофаминов при индукции апоптоза в культуре клеток мозжечка.
Нейропротекторный эффект эндогенных каннабиноидов анандамида и 2-арахидоноилглицерина при моделировании различных форм церебральной патологии был показан рядом авторов (van der Stelt et al., 2001; Baker et al., 2003; Panikashvili et al., 2001, 2005; Eljaschewitsch et al., 2006). Показано участие эндоканнабиноидов в регуляции процессов, связанных с выживаемостью нейронов, при повреждении которых в нервной ткани происходит усиление продукции эндогенных лигандов КР (Stella et al., 1997), а нейроны, генетически лишенные КР, более чувствительны к действию возбуждающих аминокислот (Marsicano et al., 2003). Однако защитный эффект ряда N-ацилдофаминов, а также метаболитов ЭК остается недостаточно изученным.
Нами исследовано защитное действие агонистов КР, относящихся к группе N-ацилдофаминов, на модели нейронального апоптоза в первичной культуре клеток-зерен мозжечка крысы (модель К+-депривации). Снижение концентрации К+ в инкубационной среде приводит к индукции апоптоза. Апоптоз является типом программированной клеточной гибели, которая лежит в основе многих физиологических и патофизиологических состояний нервной системы, в том числе, ишемии (Ramiro-Cortes, Moran, 2009). Независимо от причины, вызвавшей локальное ишемическое повреждение мозга, развивается каскад патобиохимических изменений, приводящих к необратимому повреждению нервной ткани по механизмам некроза и/или апоптоза (Гусев Е.И. с соавт., 1997).
Прижизненная микроскопия культур обнаружила, что через 20 часов после индукции
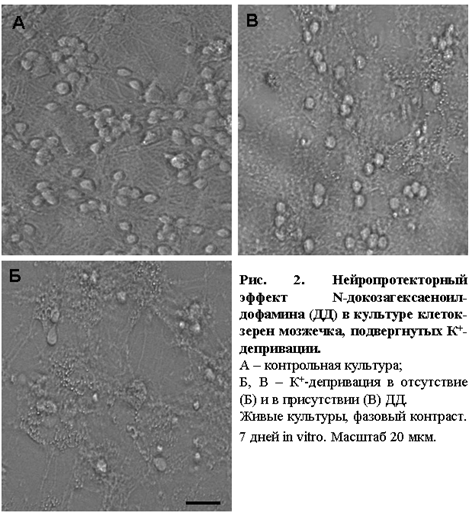
апоптоза (К+-депривации) в отсутствие N-ацилдофаминов происходит необратимая дегенерация большей части нейронов (рис. 2Б), тогда как глиальные клетки остаются неповрежденными. При инкубации культур, подвергнутых апоптозу, в присутствии N- докозагексаеноилдофамина (ДД) в них сохраняется значительное количество живых нейронов (рис. 2В).
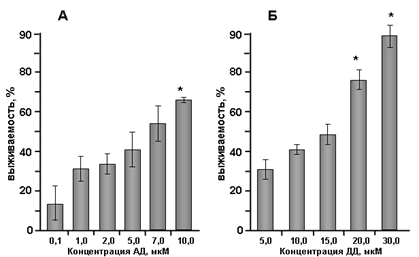
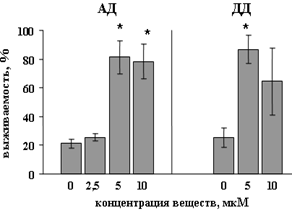
Биохимический анализ выживаемости при помощи МТТ-теста показал, что в присутствии АД и ДД жизнеспособность клеток значительно повышается (рис. 3). Так, максимальная выживаемость культур (до 65 %) под действием АД (рис. 3А) в диапазоне от 0,1 до 10 мкМа достигалась при концентрации 10 мкМ, однако дальнейшее повышение концентрации оказывало цитотоксический эффект. В присутствии ДД (в концентрации от 1 до 30 мкМ, рис. 3Б) выживаемость также дозозависимо возрастала, достигая 86 %, при этом цитотоксический эффект во всем диапазоне концентраций не наблюдался.
|
Рис. 3. Нейропротекторное действие N-ацилдофаминов N-арахидоноилдофамина (АД) (А) и N-докозагексаеноилдофамина (ДД) (Б) в культуре клеток мозжечка, подвергнутых апоптозу. МТТ-тест. n=40. *р<0,05 (по данным дисперсионного анализа) |
|
Рис. 4. Влияние антагонистов каннабиноидных рецепторов на защитный эффект N-ацилдофаминов N-арахидоноилдофамина (АД) (А) и N-докоза-гексаеноилдофамина (ДД) (Б) при апоптозе, индуцированном К+-депривацией. К+ 25 мМ - концентрация калия в нормальных условиях; К+ 5 мМ - концентрация калия, индуцирующая апоптоз; +SR1 - инкубация совместно с SR141716А; +SR2 - инкубация совместно с SR144528. n=15. *р<0,05 по сравнению с контролем (критерий Вилкоксона-Манна-Уитни), зр<0,05 по сравнению с N-ацилдофаминами (критерий Вилкоксона-Манна-Уитни). |
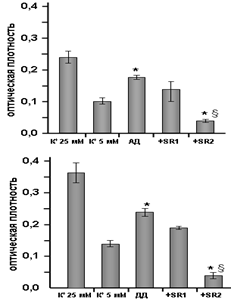
Для изучения вклада каннабиноидных рецепторов в защитные механизмы были использованы антагонисты КР1 (SR141716А) и КР2 (SR144528). SR141716А не оказывал влияния на нейропротекторный эффект, в то время как SR144528 снижал защитное действие АД более чем вдвое, а ДД - почти на 85 % (рис. 4). Поэтому основные эффекты N-ацилдофаминов могут быть связаны с активацией КР2, экспрессирующихся и на глиальных клетках, и на клетках-зернах мозжечка.
Таким образом, эти данные подтверждают предположение о важной роли взаимодействий между нейронами и клетками глии в реализации нейропротекторных свойств агонистов КР. Основываясь на полученных нами результатах, можно предположить, что КР1 и КР2 в разной степени вовлечены в защитный эффект модуляторов ЭКС, причем более значительную роль в механизмах этого эффекта играют КР2.
1.3. Нейропротекторное действие N-ацилдофаминов при цитотоксическом действии глутамата в культуре клеток новой коры.
В культурах клеток новой коры, подвергнутых цитотоксическому действию глутамата (100 мкМ, 15 мин), АД и ДД также оказывали защитный эффект (рис. 5).
Для АД максимальный эффект выживаемости (до 92 %) достигался при концентрации 5 мкМ. Присутствие ДД в той же концентрации в инкубационной среде также обеспечивало высокий показатель выживаемости клеток (рис. 5).
|
Рис. 5. Нейропротекторное действие N-арахидоноилдофамина (АД) и N-докозагексаеноилдофамина (ДД) в культуре клеток новой коры при цитотоксическом действии глутамата (100 мкМ, 15 мин). Данные МТТ-теста, приведенные в % к показателям выживаемости под действием глутамата в отсутствие каннабиноидов. Число культур n=15. *р<0,05 по сравнению с контролем (Критерий Вилкоксона-Манна-Уитни). |
1.4. Нейропротекторные свойства N-докозагексаеноилдофамина при цитотоксическом действии глутамата в органотипической культуре ткани гиппокампа.
Ряд работ по изучению нейропротекторного действия каннабиноидов и участия в нем КР был выполнен на органотипических эксплантатах гиппокампа (Khaspekov et al., 2004; Kreutz et al., 2007; Koch et al., 2011). АД, добавленный в культуральную среду срезов гиппокампа, подвергнутых токсическому повреждению N-метил-D-аспартата дозозависимо защищал зернистый слой зубчатой фасции путем активации КР1 (Grabiec et al., 2012).
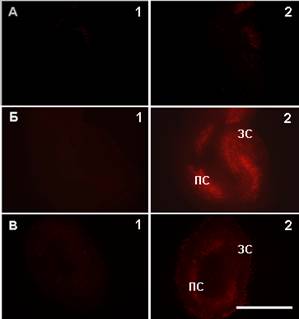
В наших экспериментах ДД в эксплантатах гиппокампа, как и в культурах клеток-зерен мозжечка и новой коры, токсического действия на нейроны не оказывал (рис. 6А). После воздействия глутамата (200 мкМ, 15 мин) через 24 часа в пирамидном слое аммонова рога и зернистом слое зубчатой фасции при окраске флуоресцентным красителем пропидий иодидом, выявлялись ядра погибших нейронов (рис. 6Б). Инкубация культур с ДД (20 мкМ) в течение 24 часов после воздействия глутамата оказывала выраженный нейропротекторный эффект (рис. 6В).
|
Рис. 6. Нейропротекторное действие N-докозагексаеноилдофамина (ДД) в органотипической культуре ткани гиппокампа при цитотоксическом действии глутамата. А - контрольный эксплантат, интервал между 1 и 2 - 24 часа; Б - эксплантат до (1) и через 24 часа после (2) 15-минутного воздействия глутамата (200 мкМ); В - эксплантат до (1) и через 24 часа после (2) 15-минутного воздействия глутамата и дальнейшей инкубации в присутствии ДД (20 мкМ). ПС - пирамидный слой аммонова рога; ЗС - зернистый слой зубчатой фасции. Окраска - пропидий иодид. 12 (1) - 13 (2) дней культивирования. Масштаб 1 мм. |
Таким образом, каннабимиметики АД и ДД дозозависимо, в пределах определенных концентраций, препятствуют повреждению культивируемых нейронов, вызванному апоптозом и цитотоксическим действием глутамата, что позволяет рассматривать эти N-ацилдофамины как потенциальные и эффективные нейропротекторы.
1.5. Модуляция спонтанной биоэлектрической активности нейронной сети первичной культуры гиппокампа глутаматом и N-ацилдофаминами.
Клетки гиппокампа, культивируемые на мультиэлектродной матрице, формируют нейронные сети, в которых в замкнутом пространстве, без воздействия экзогенных стимулирующих факторов, происходит обмен информацией между ограниченным количеством нейронов (Мухина с соавт., 2009). Функционирование таких сетей регистрируется в виде спонтанной синхронной сетевой пачечной активности (СПА), представляющей собой генерируемые одновременно несколькими нейронами последовательности потенциалов действия (спайков) небольшой продолжительности (несколько сотен миллисекунд). Сложный рисунок СПА, регистрируемый каждым из электродов мультиэлектродной матрицы, характеризуется определенным количеством пачек и частотой спайков в пачке.
В наших экспериментах, проводившихся на 14-й день in vitro, действие глутамата в диапазоне концентраций от 0,5 до 100 мкМ вызывало дозозависимое повышение уровня СПА нейронов гиппокампа с изменением паттерна активности (таблица 1). При этом происходило возрастание частоты генерации сетевых пачек импульсов при снижении количества импульсов в каждой пачке.
Таблица 1. Влияние глутамата на показатели биоэлектрической активности нейронных сетей в культуре клеток гиппокампа.
Показатель |
Концентрация глутамата, мкМ |
||||||
0 |
0,5 |
1 |
2 |
10 |
50 |
100 |
|
Количество пачек/5 мин |
16,7 3,34 |
34,8 2,88* |
59,5 3,23*,** |
61,8 2,55* |
72,0 5,52* |
60,5 2,77* |
62,4 4,12* |
Количество спайков/пачка |
2987 466 |
1655 438* |
1120 227*, ** |
1008 269* |
1104 186* |
420 86*, ** |
350 62* |
*р<0,05 - достоверное отличие от исходного уровня; **р<0,05 - достоверное отличие от предыдущего значения. Число культур n=20 (критерий Вилкоксона-Манна-Уитни).
Анализ растровых диаграмм СПА при аппликации высоких концентраций глутамата (от 10 до 100 мкМ) обнаружил возникновение относительно короткого периода молчания активности нейронов, возможно, связанного с десенситизацией глутаматных рецепторов или же длительной деполяризацией нейрональной мембраны. Кроме того, на ряде электродов при аппликации больших доз глутамата регистрировалось полное либо частичное подавление СПА. Объяснением этого эффекта может быть гиперактивация ГАМКергических интернейронов гиппокампа в нейронной сети, приводящая к подавлению активности синаптически связанных с ними пирамидных клеток. Однако преинкубация культур с антагонистом ГАМКА-рецепторов пикротоксином (2 мкМ) торможение СПА полностью не устраняло. Для детального выяснения механизмов указанных эффектов необходимы дополнительные исследования.
Действие АД в концентрации до 5 мкМ практически не влияло на параметры СПА (таблица 2), тогда как при концентрации 7 и 10 мкМ сетевой паттерн активности нейронной сети изменялся, при этом происходило постепенное снижение уровня активности СПА. При сочетанном введении глутамата (50 мкМ) и АД (10 мкМ) восстановление синхронизированной пачечной активности нейронов после периода молчания происходило быстрее, чем при действии глутамата в отсутствие АД.
Таблица 2. Влияние N-арахидоноилдофамина (АД) на показатели биоэлектрической активности нейронных сетей в культуре клеток гиппокампа.
Показатель |
Концентрация АД, мкМ |
||||||
0 |
0,5 |
1 |
2 |
5 |
7 |
10 |
|
Количество пачек/5 мин |
18,5 3,55 |
17,9 4,37 |
18,5 5,23 |
16,4 3,67 |
15,7 3,47 |
15 5,12 |
16 2,85 |
Количество спайков/пачка |
2005 423 |
1675 328 |
1358 312 |
1469 387 |
1623 233 |
1050 148*, ** |
645 134* |
*р<0,05 - достоверное отличие от исходного уровня; **р<0,05 - достоверное отличие от предыдущего значения. Число культур n=10 (критерий Вилкоксона-Манна-Уитни).
ДД в концентрациях до 20 мкМ в отсутствие глутамата не оказывал заметного влияния на СПА в нейронной сети. Результаты совместного действия глутамата (50 мкМ) и ДД (20 мкМ) приведены в таблице 3. Видно, что под влиянием ДД происходило изменение сетевых параметров СПА за счет снижения частоты следования пачек в минуту и возрастания количества спайков в пачке (табл. 3).
Таблица 3. Влияние глутамата и N-докозагексаеноилдофамина (ДД) на показатели биоэлектрической активности нейронных сетей в культуре клеток гиппокампа.
Показатель |
Исходная культура |
Глутамат (50 мкМ) |
Глутамат (50 мкМ) + ДД (20 мкМ) |
Количество пачек/мин |
8,3 1,64 |
30,04 4,01* |
19,9 2,62* |
Количество спайков/пачка |
467,2 50,2 |
173,3 33,7* |
339 68,7* |
*р<0,05 - достоверное отличие от предыдущего значения; n=10 (критерий Вилкоксона)
Эффекты от добавления N-ацилдофаминов в инкубационную среду после воздействия глутамата (50 мкМ), возможно, связаны с нормализацией синаптической передачи и частичным устранением повреждающего действия больших доз глутамата.
Полученные данные могут свидетельствовать о способности N-ацилдофаминовых аналогов эндоканнабиноидов модулировать спонтанную активность нейронной сети, измененную глутаматным воздействием, и таким образом препятствовать его негативному влиянию на функциональное состояние нейронов.
1.6. Нейропротекторное действие N-ацилдофаминов при их совместном применении с ингибиторами гидролазы амидов жирных кислот.
Серьезным затруднением в реализации защитного действия эндоканнабиноидов является их довольно быстрый гидролиз в клетках и тканях (Di Marzo, 2008). Поэтому мы предположили, что совместное применение каннабиноидов и ингибиторов основного фермента их гидролиза, гидролазы амидов жирных кислот, позволит потенциировать и пролонгировать нейропротекторное действие агонистов КР и таким образом снизить их действующую концентрацию.
Нами были исследованы защитные свойства двухкомпонентных систем, в которых первым компонентом является лиганд КР, а второй компонент представлен ингибитором ГАЖК. Ранее было установлено, что N-арахидоноилсеротонин (АС) является селективным ингибитором ГАЖК (Bisogno et al., 1998). Было обнаружено, что наиболее активным соединением, аналогом АС, является N-эйкозапентаеноилсеротонин (ЭС) (Генрихс с соавт., 2010). Поэтому в качестве первого компонента исследуемого комплекса веществ нами были выбраны N-арахидоноилдофамин и N-докозагексаеноилдофамин, а второго - АС и его более активный аналог ЭС.
Сами по себе ингибиторы ГАЖК в культуре клеток-зерен мозжечка защитного эффекта не оказывали. При использовании АС в качестве второго компонента в комплексе с АД и ДД выраженного усиления защитного эффекта не происходило (таблица 4), что, возможно, связано с меньшей активностью АС в отношении ГАЖК.
Таблица 4. Влияние ингибиторов ГАЖК на защитный эффект N-ацилдофаминов в культуре клеток мозжечка.
Каннабиноид |
Без ингибитора |
ЭС 10 мкМ |
АС 10 мкМ |
АД, 5 мкМ |
40 3,2 |
80,8 4,3* |
43 5,6 |
ДД, 5 мкМ |
27 4,4 |
94 3,1* |
32,1 3,7 |
Указан процент выживаемости культур по сравнению с контрольными, подвергнутыми К+-депривации без модуляторов ЭКС. *p<0,05 по сравнению с защитным действием N-ацилдофаминов в отсутствие ингибиторов ГАЖК. Данные МТТ-теста. n=10 (критерий Вилкоксона-Манна-Уитни).
Использование ЭС в качестве второго компонента достоверно усиливало защитный эффект АД, взятого в субмаксимальной концентрации (5 мкМ). Максимальное нейропротекторное действие для комплекса АД с ЭС при концентрации последнего 10 мкМ составило 80 %. Комплекс ДД с ЭС также значительно повышал жизнеспособность культур по сравнению с контролем, а увеличение концентрации ЭС до 10 мкМ позволило достигнуть максимальной (до 94 %) выживаемости клеток (таблица 4).
Нейропротекторный эффект АД в субмаксимальной концентрации (2,5 мкМ) в комплексе с ЭС в диапазоне концентраций от 0,5 до 5 мкМ исследован нами при цитотоксическом действии глутамата (100 мкМ) в культуре клеток новой коры. Максимальное защитное действие этого комплекса (выживаемость до 79 %) было достигнуто при концентрации ЭС 1,25 мкМ (рис. 7). Дальнейшее увеличение концентрации ЭС приводило к снижению защитного эффекта.
*аа а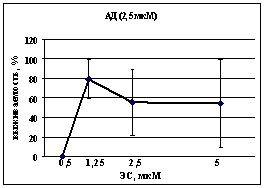 |
Рис. 7. Модулирующее влияние различных концентраций ЭС на защитный эффект АД (2,5 мкМ) в культуре клеток новой коры, подвергнутой цитотоксическому действию глутамата (100 мкМ). Указан процент выживаемости культур по сравнению с контрольными, подвергнутыми действию глутамата без модуляторов эндогенной каннабиноидной системы. Данные МТТ-теста. n=5. *p<0,05 (критерий Вилкоксона-Манна-Уитни) |
Ранее было показано, что N-ацилдофамины являются антиоксидантами и защищают нейроны в условиях окислительного стресса in vitro (Бобров с соавт., 2006). В работе по изучению влияния N-ацилдофаминов на микроциркуляцию обнаружена их способность усиливать локальный кровоток в головном мозге крыс, а также оказывать антиагрегационное действие (Васильева с соавт., 2002). Кроме того, N-ацилсеротонины, помимо их ингибиторных свойств в отношении ГАЖК, способны эффективно блокировать агрегацию тромбоцитов человека, стимулированную арахидоновой кислотой, АДФ и адреналином (Васильева с соавт., 2002). Приведенные данные публикаций указывают на то, что нейролипины могут выступать как нейропротекторы на разных этапах развития ишемических повреждений головного мозга.
Таким образом, полученные нами результаты в сопоставлении с данными литературы позволяют заключить, что использование субмаксимальных концентраций каннабиноидов в сочетании с ингибиторами ГАЖК приводит к значительному усилению защитного эффекта. Важно отметить, что исследованные ингибиторы не оказывают собственного защитного действия и в пределах использованных концентраций не токсичны. Поэтому использованная нами комбинация нейролипинов позволяет значительно снизить действующую концентрацию N-ацилдофаминов и, следовательно, уменьшить их возможный побочный эффект без снижения эффективности нейропротекции.
1.7. Нейропротекторные свойства метаболита эндогенного каннабиноида анандамида простамида Е2.
Известно, что некоторые ЭК, такие как анандамид, могут трансформироваться циклооксигеназами в окисленные производные (этаноламиды простагландина или простамиды) (Woodward et al., 2008), которые, вероятно, имеют собственный спектр биологической активности и проявляют защитные свойства. Однако участие в нейрозащитных процессах циклооксигеназных метаболитов анандамида при различных патологических состояниях практически не изучалось. В наших исследованиях использован циклооксигеназный метаболит анандамида простамид Е2 (ПАЕ2).
Как анандамид, так и ПАЕ2 нейроцитотоксического действия на клетки не оказывали. Однако, в то время как анандамид в концентрациях до 10 мкМ в условиях К+-депривации не проявлял защитных свойств (выживаемость клеток была такой же, как в негативном контроле), ПАЕ2 дозозависимо оказывал нейропротекторный эффект, максимальный в концентрации 1 мкМ, повышая выживаемость культивируемых клеток мозжечка до 88 % (таблица 5).
Таблица 5. Выживаемость нейронов в культурах клеток мозжечка, подвергнутых апоптозу, в присутствии анандамида и простамида Е2 (в %).
Соединение |
Концентрация, мкМ |
|||
0 |
0,1 |
0,5 |
1,0 |
|
Анандамид |
44,9 2,3 |
45,2 1,1 |
45 0,6 |
47,4 3,0 |
ПАЕ2 |
46,2 1,3 |
61,1 8,6* |
79,3 4,9* |
88,4 2,8* |
Данные МТТ-теста. n=10. *р<0,05 - достоверное различие по сравнению с контролем (критерий Вилкоксона-Манна-Уитни).
Таким образом, циклооксигеназная трансформация анандамида способна приводить к образованию производных с новым спектром биологической активности и выраженным нейропротекторным действием.
2. Нейропротекторное действие модуляторов ЭКС in vivo.
2.1. Патогистологическое исследование.
В исследовании были использованы N-ацилдофамин ДД и его комбинация с ЭС (ингибитором ГАЖК). Сразу после индукции фототромбоза участка теменно-височной области новой коры, а далее однократно в течение 4-х суток крысам внутрибрюшинно вводили ДД в дозе 5 мг/кг, либо в концентрации 2,5 мг/кг в сочетании с ЭС (5 мг/кг). Контрольным крысам после ФТ по той же схеме вводили физиологический раствор. При визуальном осмотре очаговое поражение мозга контрольной группы животных выглядело значительно более выраженным, чем у животных, которым вводились модуляторы ЭКС.
При гистологическом анализе срезов мозга после индукции ФТ в отсутствие модуляторов ЭКС в области ишемического повреждения выявлялся участок тотального некроза, окруженный зоной т.н. пенумбры (от латинского УpenumbraФ - полутень), в ткани которой обнаруживался перицеллюлярный и периваскулярный отек с диффузным смещением клеток и содержалисьа нейроны с начальными изменениями структуры, характерными для данной стадии постишемического периода, такими как гиперхромность, сморщивание тел нейронов, гомогенизация цитоплазмы, изменение конфигурации ядра. После индукции ФТ с введением модуляторов ЭКС в зоне пенумбры преобладали нейроны с неизмененной структурой, а периваскулярный отек был выражен слабее.
Объем ишемического очага при введении ДД или ДД в сочетании с ЭС не достигал объема, наблюдаемого при индукции ФТ в отсутствие модуляторов ЭКС (рис. 8).
* а*аа а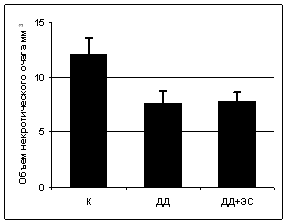 |
Рис. 8. Защитный эффект N-докозагекса-еноилдофамина (5 мг/кг, ДД) и его комбинации (2,5 мг/кг) с N-эйкозапента-еноилдофамином (5 мг/кг, ДД+ЭС) при фототромбозе коры головного мозга крыс. К - в отсутствие модуляторов ЭКС. *р<0,05 по отношению к контролю. Данные морфометрического анализа. Число животных, использованных в каждом эксперименте n=15 (критерий Вилкоксона-Манна-Уитни). |
Таким образом, модуляторы ЭКС, использованные нами при фокальной ишемии коры головного мозга крыс, индуцированной фототромбозом, обладают выраженным защитным действием. Важно отметить, что степень нейропротекторного эффекта ДД в дозе 5 мг/кг без участия ЭС оказалась сходной с наблюдавшейся нами при значительно более низкой дозе ДД (2,5 мг/кг) при его использовании в сочетании с ЭС. Это подтверждает обнаруженную нами в экспериментах in vitro (см. выше) возможность снижения эффективной концентрации N-ацилдофаминов при их сочетанном применении с ингибиторами ГАЖК.
2.2. Электронно-микроскопическое исследование.
В пенумбре на 4-е сутки после ФТ III-V корковые слои содержат нейроны, измененные как по светлому, так и по темному типу. Такая вариабельность ультраструктуры нейронов может быть обусловлена, вероятно, их индивидуальной чувствительностью и различным функциональным состоянием при локальном нарушении кровообращения в зоне повреждения (Enright et al., 2007). Появление электронно-светлых нейронов связано, скорее всего, с их отеком, тогда как гиперхроматоз может быть сигналом к необратимому развитию отсроченной гибели по типу апоптоза, первичными признаками которого, наблюдавшимися в наших экспериментах, является наличие в ядрах клампированного хроматина и частичная дезагрегация полисом при нормальной ультраструктуре органелл (Hossmann et al., 2001).
Апоптотические изменения ультраструктуры нейронов после ФТ при введении ДД не обнаруживались, что является важным свидетельством его нейропротекторного эффекта. Кроме того, появление в цитоплазме ишемизированных нейронов органелл с признаками восстановления интактной ультраструктуры и энергизованных митохондрий указывает на активацию в клетках репаративных процессов.
Уменьшение количества шипиков, синапсов и синаптических пузырьков, наблюдавшееся нами в нейропиле после ФТ, согласуется с данными литературы, указывающими на то, что такие изменения в нейронах пенумбры неокортекса у крыс происходят в течение первых четырех суток после инсульта (Ito et al., 2007), причем наличие удлиненных шипиков и синапсов на них связывают с появлением аберрантной синаптической передачи. Предполагается, что изменение числа и длины шипиков является ранним адаптивным ответом потенциально чувствительных нейронов в пенумбре (Brown et al., 2008) и связано с постишемическим распространением депрессивно-подобной деполяризации и потерей синапсов. Нами обнаружено, что под воздействием ДД ультраструктура нейропиля нормализуется, а именно просветляется матрикс аксонов, дендритов и шипиков, причем число шипиков, а также их синаптических контактов с аксонными терминалями в поле зрения возрастает. Характерной особенностью нейропиля после введения ДД является появление крупных шипиков с хорошо выраженным шипиковым аппаратом, а также множественных сложно организованных и перфорированных синапсов, что указывает на повышение эффективности синаптической передачи (Marrone and Pettit, 2002).
Полученные данные свидетельствуют о том, что N-ацилдофамины при фокальной ишемии могут способствовать репаративным процессам в пенумбре, происходящим в результате пластических перестроек ультраструктуры нейронов и межнейронных связей.
ВЫВОДЫ
- N-ацилдофаминовые аналоги эндоканнабиноидов N-арахидоноилдофамин и N-докозагексаеноилдофамин оказывают нейропротекторный эффект, повышая жизнеспособность клеток, при индукции апоптоза и цитотоксическом действии глутамата in vitro.
- Защитное действие исследованных N-ацилдофаминов N-арахидоноилдофамина и N-докозагексаеноилдофамина при индукции апоптоза in vitro опосредуется каннабиноидными рецепторами 2-го типа.
- N-арахидоноилдофамин и N-докозагексаеноилдофамин нормализуют вызванные глутаматом изменения биоэлектрической активности нейронов в функциональной нейронной сети, сформированной клетками гиппокампа на мультиэлектродной матрице.
- Ингибирование гидролиза N-ацилдофаминов N-арахидоноилсеротонином и N-эйкозапентаеноилсеротонином приводит к повышению эффективности их защитного действия, увеличивая жизнеспособность нейронов.
- Модуляторы эндогенной каннабиноидной системы препятствуют увеличению объема ишемического очага, активируют репаративные процессы в пенумбре, происходящие в результате пластических перестроек ультраструктуры нейронов и межнейронных связей, и, таким образом, оказывают нейропротекторный эффект при моделировании фокальной ишемии головного мозга крыс, индуцированной фототромбозом кровеносных сосудов новой коры in vivo.
- Полученные результаты могут служить экспериментальным обоснованием возможности разработки на основе модуляторов эндогенной каннабиноидной системы новых терапевтических подходов в целях фармакологической коррекции повреждений нейронов головного мозга при церебральной ишемии.
Практические рекомендации. Полученные результаты можно рекомендовать к их практическому использованию в учебных материалах для студентов медицинских и биологических факультетов вузов. Данные исследования дают основу для их практического применения при дальнейших экспериментальных разработках, направленных на получение новых нейропротекторных фармацевтических соединений на основе модуляторов эндогенной каннабиноидной системы.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- Генрихс, Е.Е. Модуляторы эндогенной каннабиноидной системы как нейропротекторы / Е.Е. Генрихс, М.Ю. Бобров, Е.Л. Андрианова, Н.М. Грецкая, А.А. Лыжин, А.В. Блаженова, Л.Е. Фрумкина, В.В. Безуглов, Л.Г. Хаспеков // Анналы клинической и экспериментальной неврологии. - 2010. - № 4 - С. 37-42.
- Генрихс, Е.Е. Нейропротекторные свойства модуляторов эндогенной каннабиноиднойа системы in vivo и in vitro / Е.Е. Генрихс, И.В. Барсков, М.Ю. Бобров, Е.Л. Андрианова, А.А. Лыжин, Л.Г. Хаспеков // Сборник трудов Всероссийской конференции Современные направления в исследовании функциональной межполушарной асимметрии и пластичности мозга. Москва. - 2010. - С. 338-342.
- Генрихс, Е.Е. Модуляция спонтанной биоэлектрической активности нейронной сети первичной культуры гиппокампа глутаматом и N-арахидоноилдофамином / С.А. Коротченко, М.В. Ведунова, Е.А. Корягина, М.Ю. Бобров, Е.Е. Генрихс, Л.Г. Хаспеков, И.В. Мухина // Сборник трудов Всероссийской конференции Современные направления в исследовании функциональной межполушарной асимметрии и пластичности мозга. Москва. - 2010. - С. 390-394.
- Генрихс, Е.Е. Ультраструктурное исследование нейропротекторного действия каннабиноида N-докозагексаеноилдофамина при фокальной ишемии коры головного мозга крыс / Л.Е. Фрумкина, Е.Е. Генрихс, И.В. Барсков, М.Ю. Бобров, М.В. Оборина, Е.Л. Андрианова, Л.Г. Хаспеков // Сборник трудов Всероссийской конференции Современные направления в исследовании функциональной межполушарной асимметрии и пластичности мозга. Москва. - 2010. - С. 507-511.
- Генрихс, Е.Е.а Эндоканнабиноиды и их производные как нейропротекторы / Е.Е. Генрихс, Е.Л. Андрианова, М.Ю. Бобров // Материалы IV Международной научной конференции молодых ученых-медиков. 25-26 февраля 2010 года. / Под редакцией В.А. Лазаренко - Курск: ГОУ ВПО КГМУ Росздрава. - 2010. - Т. 1 - С. 264-266.
- Генрихс, Е.Е. Нейропротекторное действие эндоканнабиноидов и их производных / Е.Е. Генрихс, Е.Л. Андрианова, М.Ю. Бобров, А.А. Лыжин,а Н.М. Грецкая, В.В. Безуглов, Л.Г. Хаспеков // Сборник трудов 6-го Международного междисциплинарного конгресса Нейронаука для медицины и психологии. Судак, Крым, Украина, 5-15 июня 2010 г. / Под редакцией Е.В. Лосевой, Н.А. Логиновой, В.В. Синельниковой - Москва, - 2010. - С. 91-92.
- Генрихс, Е.Е. Действие анандамида и его циклооксигеназного производного, простамида Е2, на нормальные и трансформированные нервные клетки in vitro / Е.Л. Андрианова, Е.Е. Генрихс, М.Ю. Бобров, А.А. Лыжин, Н.М. Грецкая, Л.Е. Фрумкина, Л.Г. Хаспеков, В.В. Безуглов // Бюллетень экспериментальной биологии и медицины. - 2011. - № 151 - С. 36-38.
- Генрихс, Е.Е. Нейропротекторный эффект модуляторов эндогенной каннабиноидной системы при экспериментальной церебральной ишемии / М.Ю. Бобров, Е.Е. Генрихс, И.В. Барсков, А.А. Лыжин, Л.Е. Фрумкина, Е.Л. Андрианова, М.В. Оборина, Л.Г. Хаспеков // Сборник статей Международной конференции Рецепторы и внутриклеточная сигнализация / Под редакцией В.П. Зинченко, С.С. Колесникова, А.В. Бережнова. Пущино. - 2011. - Т. 1 - С. 94-98.
- Genrikhs, E.E. Modulation of spontaneous network activity in hippocampal cell culture by glutamate and N-arachidonoyl dopamine / L.G. Khaspekov, S.A Korotchenko, M.V. Vedunova, E.A. Koryagina, M.Yu. Bobrov, E.E. Genrikhs, I.V. Mukhina // Proc. of the III International Symposium "Topical Problems of Biophotonics - 2011". Nizhny Novgorod. - 2011. - P. 232-233.
Список использованных сокращений.
АД - N-арахидоноилдофамин
АДФ Цаденозиндифосфат
Анандамид - N-арахидоноилэтаноламин
АС - N-арахидоноилсеротонин
ГАМК - гамма-аминомасляная кислота
ГАЖК - гидролаза амидов жирных кислот
ДД - N-докозагексаеноилдофамин
КР - каннабиноидный рецептор
КР1 - каннабиноидные рецепторы 1-го типа
КР2 - каннабиноидные рецепторы 2-го типа
ПАЕ2 - простамид Е2
СПА - сетевая пачечная активность
ФТ - фототромбоз
ЭК - эндоканнабиноиды
ЭКС - эндогенная каннабиноидная система
ЭС - N-эйкозапентаеноилсеротонин
Авторефераты по темам >>
Разные специальности - [часть 1] [часть 2]